КТР по неделям беременности: что такое, таблица
Копчиково-теменной размер или КТР по неделям беременности, возможно, измерить только на первом обязательном скрининге. Этот диагностический показатель помогает не только установить срок гестации, но и предоставляет возможность судить о степени развития малыша внутри утробы.Что означает КТР плода на узи при беременности?
КТР на узи при беременности – это основной параметр, определяющий величину и размер эмбриона. Измерения наиболее достоверны на этапе гестации от 10 до 16 недель. Раньше этого периода плод еще слишком мал для подобных измерений, позднее диагностический метод уже не такой информативный.Понимание КТР при беременности дает возможность определить точный срок вынашивания. Благодаря копчико-теменному размеру врачи оценивают динамические показатели роста, рассчитывают вероятность формирования крупного плода, а также сравниваются риски по задержке внутриутробного развития.
На этапе ультразвукового скрининга специалисты проводят продольные замеры, которые не включают в себя конечности эмбриона. Не редко применяется трехкратное измерение, когда в течение 1 исследования делают три замера в период максимального разгибания младенца, но берут во внимание только усредненные размеры.
Не редко применяется трехкратное измерение, когда в течение 1 исследования делают три замера в период максимального разгибания младенца, но берут во внимание только усредненные размеры.
Срок беременности определяется в соответствии с некоторыми правилами:
- показатель < 10 мм + 6;
- > 10 мм + 6,5.
Таблица нормы ктр по неделям беременности
Размер ктр по неделям беременности с течением времени возрастает, так как плод растет и развивается. В случае проведения УЗИ несколько раз, то копчико-теменная длина изучается по динамическим свойствам, что позволяет исключить различные патологии.
Таблица нормы ктр по неделям беременности:
| КРТ, мм | Недели + дни | КРТ, мм | Недели + дни | КРТ, мм | Недели + дни |
|---|---|---|---|---|---|
| 7 | 6+3 | 32 | 10 | 57 | 12+2 |
| 8 | 6+4 | 33 | 10+1 | 58 | 12+3 |
| 9 | 6+6 | 34 | 10+2 | 59 | 12+3 |
| 10 | 7 | 35 | 10+2 | 60 | 12+4 |
| 11 | 7+2 | 36 | 10+3 | 61 | 12+4 |
| 12 | 7+3 | 37 | 10+4 | 62 | 12+5 |
| 13 | 7+4 | 38 | 10+5 | 63 | 12+5 |
| 14 | 7+5 | 39 | 10+6 | 64 | 12+6 |
| 15 | 7+6 | 40 | 11 | 65 | 12+6 |
| 16 | 8 | 41 | 11 | 66 | 13 |
| 17 | 8+1 | 42 | 11+1 | 67 | 13 |
| 18 | 8+2 | 43 | 11+2 | 68 | 13+1 |
| 19 | 8+3 | 44 | 11+2 | 69 | 13+1 |
| 20 | 8+4 | 45 | 11+3 | 70 | 13+2 |
| 21 | 8+5 | 46 | 11+3 | 71 | 13+2 |
| 22 | 8+6 | 47 | 11+4 | 72 | 13+3 |
| 23 | 9 | 48 | 11+5 | 73 | 13+3 |
| 24 | 9+1 | 49 | 11+5 | 74 | 13+4 |
| 25 | 9+2 | 50 | 11+6 | 75 | 13+4 |
| 26 | 9+3 | 51 | 11+6 | 76 | 13+5 |
| 27 | 9+3 | 52 | 12 | 77 | 13+5 |
| 28 | 9+4 | 53 | 12+1 | 78 | 13+5 |
| 29 | 9+5 | 54 | 12+1 | 79 | 13+6 |
| 30 | 9+6 | 55 | 12+2 | 80 | 13+6 |
| 30 | 10 | 56 | - | - | - |
Если норма КТР больше срока, то зачастую это указывает на раннюю диагностику крупного плода.
Отклонения КТР в меньшую сторону
Чаще всего, при значительно сниженных показателях предусматривают неверно рассчитанный акушерский срок вынашивания. Овуляционный период и оплодотворение яйцеклетки, в этом случае, наступили позднее. Чтобы перепроверить данные назначают повторное обследование УЗИ по истечению примерно недели.К другим отклонениям относятся:
- остановившаяся в развитии беременность, когда эмбрион погибает внутри утробы, вследствие определенных патологических процессов. Диагноз ставится по отсутствию сердечных сокращений и двигательной активности. Ситуация требует незамедлительного оперативного вмешательства, чтобы не спровоцировать осложнения.
-
недостаточное продуцирование прогестерона или других гормонов может закончиться выкидышем. Назначается дополнительное УЗ-исследование и медикаментозная терапия с целью нормализации гормонального фона.

- наличие инфекционных заболеваний также обуславливается нарушением роста и формированием ребенка. Проводятся дополнительные анализы, предусматривается лечение инфекции.
- подозрение на нарушения генетического характера, связанные с различными синдромами, подтверждаются с помощью анализов 3-х маркеров – скрининг, ХГЧ и протеина-А.

что это такое, какая норма, как измеряется показатель?
КТР плода – важный параметр, позволяющий отслеживать внутриутробное развитие ребенка и своевременно принимать меры для устранения возникающих нарушений. Установление копчико-теменного размера осуществляется посредством проведения УЗИ. Расшифровка результата исследования производится с помощью специальной таблицы, в которой указаны нормальные значения этого показателя для каждой недели гестации. Каким в идеале должен быть КТР плода по неделям? Что означает отклонение от нормальных значений?
Каким в идеале должен быть КТР плода по неделям? Что означает отклонение от нормальных значений?
Содержание
- 1. Что такое показатель КТР, как измеряется параметр?
- 2. Норма копчико-теменного размера плода в зависимости от недели беременности (таблица)
- 3. Причины отклонений
- 3.1. Замершая беременность
- 3.2. Сбои в гормональном фоне женщины
- 3.3. Заболевания инфекционного характера
- 3.4. Патологии матки и генетические болезни
- 4. Что делать в случае отклонений?
- 5. Вопросы читателей и ответы эксперта
Что такое показатель КТР, как измеряется параметр?
Эта аббревиатура расшифровывается, как «копчико-теменной размер». В акушерстве и гинекологии так называют длину прямой линии от наивысшей точки на темечке ребенка до максимально выступающей области копчика. Измерение производится в миллиметрах. Данный параметр является одним из основных показателей в процессе УЗИ на ранних сроках беременности. Этот показатель помогает установить, соответствуют ли размеры плода нормам, а также с большой точностью определить срок гестации. На копчико-теменной размер не влияют такие факторы, как пол ребенка, расовая принадлежность, цвет кожи, экологические условия, какие-либо индивидуальные особенности.
Этот показатель помогает установить, соответствуют ли размеры плода нормам, а также с большой точностью определить срок гестации. На копчико-теменной размер не влияют такие факторы, как пол ребенка, расовая принадлежность, цвет кожи, экологические условия, какие-либо индивидуальные особенности.
К определению копчико-теменного размера приступают на 6 акушерской неделе, то есть через 1 месяц после оплодотворения яйцеклетки. Именно с этого момента уже можно зафиксировать сердцебиение крохи. Замеры на 1, 2, 3, 4, 5 неделе с момента зачатия неинформативны из-за слишком малых параметров эмбриона.
По прошествии 12-13, в крайнем случае 14 недель гестации, КТР уже не измеряют, поскольку к этому сроку размеры плода уже настолько велики, что ультразвуковой датчик не может охватить его тело в полный рост. По этой причине исследование на 16-17 неделе беременности будет малоинформативным. После 14-недельного срока беременности во время процедуры УЗИ внимание уделяется другим параметрам. Наиболее точно КТР можно определить на сроке до 10 недель.
Измерение КТР с помощью УЗИ осуществляется по следующей схеме:
- Процедура проводится в сагиттальной проекции, когда туловище плода визуально делится на две равные части. При этом измерительная линия проходит строго от темени до копчика.
- Замер копчико-теменной протяженности плода производится только при отсутствии какой-либо двигательной активности эмбриона и пациентки.
- Полученное значение соизмеряется с нормами КТР. В качестве погрешности допускается 3-4 дня в большую или меньшую сторону.
- В медицинское заключение заносят данные о том, во сколько недель гестации проводилось исследование (рекомендуется делать это на 10-11 неделе), каково значение КТР плода в миллиметрах, соответствует ли оно норме, а также сведения о частоте сердцебиения и состоянии хориона.
Норма копчико-теменного размера плода в зависимости от недели беременности (таблица)
На ранних сроках гестации при отсутствии каких-либо отклонений все малыши развиваются примерно одинаковыми темпами. По этой причине на протяжении первых 5-6 недель не определены нижние значения нормы. По мере развития плода возможны отклонения от стандартных величин (верхний и нижний пороги). Нормальные показатели КТР и допустимые отклонения указаны в таблице.
По этой причине на протяжении первых 5-6 недель не определены нижние значения нормы. По мере развития плода возможны отклонения от стандартных величин (верхний и нижний пороги). Нормальные показатели КТР и допустимые отклонения указаны в таблице.
| Срок гестации | Значение КТР, мм | |||
| Недели | Дни | Низкий порог | Среднее | Верхний порог |
| 5 | 0 | Не определяется | 1,2 | 4,3 |
| 1 | 1,4 | 4,6 | ||
| 2 | 1,7 | 4,9 | ||
| 3 | 2 | 5,3 | ||
| 4 | 2,3 | 5,7 | ||
| 5 | 2,7 | 6,1 | ||
| 6 | 3,1 | 6,6 | ||
| 6 | 0 | 3,5 | 7,1 | |
| 1 | 0,4 | 4 | 7,7 | |
| 2 | 0,8 | 4,6 | 8,3 | |
| 3 | 1,3 | 5,1 | 8,9 | |
| 4 | 1,9 | 5,8 | 9,6 | |
| 5 | 2,5 | 6,4 | 10,4 | |
| 6 | 3,1 | 7,1 | 11,1 | |
| 7 | 0 | 3,8 | 7,9 | 11,9 |
| 1 | 4,5 | 8,7 | 12,8 | |
| 2 | 5,3 | 9,5 | 13,7 | |
| 3 | 6,1 | 10,4 | 14,7 | |
| 4 | 6,9 | 11,3 | 15,6 | |
| 5 | 7,8 | 12,2 | 16,6 | |
| 6 | 8,7 | 13,2 | 17,7 | |
| 8 | 0 | 9,7 | 14,2 | 18,8 |
| 1 | 10,7 | 15,3 | 20 | |
| 2 | 11,7 | 16,4 | 21,1 | |
| 3 | 12,8 | 17,5 | 22,3 | |
| 4 | 13,9 | 18,6 | 23,5 | |
| 5 | 15 | 19,9 | 24,7 | |
| 6 | 16,2 | 21,1 | 26,1 | |
| 9 | 0 | 16,3 | 22 | 27 |
| 1 | 17 | 23 | 29,1 | |
| 2 | 18,1 | 24,2 | 30 | |
| 3 | 19 | 25 | 31 | |
| 4 | 20,2 | 26,1 | 32 | |
| 5 | 21 | 27 | 34,1 | |
| 6 | 22,1 | 29 | 36 | |
| 10 | 0 | 24,1 | 31,1 | 38 |
| 1 | 25,3 | 33,1 | 41 | |
| 2 | 26 | 34 | 42 | |
| 3 | 27,1 | 35 | 43,2 | |
| 4 | 29 | 37,1 | 45 | |
| 5 | 31 | 39,3 | 47,2 | |
| 6 | 33 | 41,1 | 49 | |
| 11 | 0 | 34 | 42,2 | 50,1 |
| 1 | 35,1 | 43 | 51 | |
| 2 | 36 | 44,2 | 52,1 | |
| 3 | 37 | 45,3 | 54,1 | |
| 4 | 38,1 | 47 | 56 | |
| 5 | 39 | 48,2 | 57,1 | |
| 6 | 40,1 | 49 | 58,3 | |
| 12 | 0 | 42 | 51,3 | 59 |
| 1 | 44 | 53 | 62 | |
| 2 | 45,1 | 55 | 65,1 | |
| 3 | 47 | 57,1 | 67,1 | |
| 4 | 49,1 | 59,3 | 69,2 | |
| 5 | 50 | 61,2 | 72 | |
| 6 | 51 | 62 | 73,4 | |
| 13 | 0 | 52,1 | 63 | 75 |
| 1 | 53,1 | 65,2 | 77 | |
| 2 | 54 | 66 | 78,2 | |
| 3 | 56 | 68,1 | 80 | |
| 4 | 58 | 70 | 82,1 | |
| 5 | 59,2 | 72 | 85,2 | |
| 6 | 61 | 74 | 87,2 | |
| 14 | 0 | 63 | 76 | 89 |
Причины отклонений
В любой момент, и на 10, и на 13 неделе гестации у плода могут быть выявлены значительные отклонения КТР от нормальных значений. Если, например, на 12 неделе расстояние от темени до копчика у плода составляет 62 мм, это говорит о том, что он просто имеет крупные размеры. Если на 14-недельном сроке эта величина равна 55 мм, причиной этого явления могут быть естественные и патологические факторы.
Если, например, на 12 неделе расстояние от темени до копчика у плода составляет 62 мм, это говорит о том, что он просто имеет крупные размеры. Если на 14-недельном сроке эта величина равна 55 мм, причиной этого явления могут быть естественные и патологические факторы.
Отклонения КТР часто возникают из-за ошибки на 2-3 дня в определении акушерского срока гестации, а значит, не требует беспокойства. В ряде случаев, если этот показатель сильно разнится с данными таблицы КТР, речь может идти о какой-либо патологии.
Замершая беременность
Если данный параметр значительно отстает от нормальных значений, например, при норме 20 мм на 9 неделе гестации КТР ребенка составляет всего 12 или 14 мм, речь может идти о замирании плода. Для исключения этой патологии проверяют частоту сердцебиения и двигательную активность плода. Однако такие параметры определяются только у эмбриона, чей КТР больше 6 мм.
Если расстояние от копчика до темени ребенка превышает эту величину, а сердечные сокращения отсутствуют, проводится повторное исследование через 5-7 дней или экстренные меры по выскабливанию маточной полости и извлечению плода. Малейшее промедление в данном случае может привести к опасным, а порой и трагичным последствиям.
Малейшее промедление в данном случае может привести к опасным, а порой и трагичным последствиям.
Сбои в гормональном фоне женщины
Если на 8 неделе гестации КТР 7-8 мм, в то время как на этом сроке его величина должна составлять 17 мм, врач может предположить, что в гормональном фоне пациентке произошли нарушения. Речь идет о дефиците прогестерона, уменьшение уровня которого неизбежно во время беременности. В данной ситуации при отсутствии грамотного назначения заместительной терапии высок риск самопроизвольного прерывания беременности на 2-3 неделе гестации и более позднем сроке. Для исключения нарушения баланса гормонов используются лабораторные способы диагностики.
Заболевания инфекционного характера
При наличии в женском организме какого-либо инфекционного процесса КТР у плода может быть снижен. Например, на 9-недельном сроке гестации этот параметр составляет 16-17 мм или на 11-12 неделе беременности он равен 60 мм, что значительно ниже нормы. Чаще всего к этому приводят инфекции половых органов (хламидиоз, микоплазмоз)..jpg) Для их излечения прибегают к антибактериальным препаратам.
Для их излечения прибегают к антибактериальным препаратам.
Патологии матки и генетические болезни
Если в процессе ультразвукового исследования, проводимого в 10-11 недель, было выявлено, что расстояние от темени до копчика у эмбриона 13-16 мм, причиной такого отклонения могут быть заболевания матки. К ним относят миому, нарушения, возникшие в результате искусственного или самопроизвольного прерывания беременности в анамнезе.
Помимо этого, к понижению данного показателя могут приводить генетические патологии. В этом случае прибегают к биохимическому обследованию, проводят амниоцентез или кордоцентез.
Что делать в случае отклонений?
При патологическом отклонении от нормы показателя КТР врачи принимают меры к сохранению беременности. Методы терапии зависят от причины, вызвавшей такое явление. В остальных случаях достаточно:
- выполнять рекомендации акушера-гинеколога;
- почаще гулять на свежем воздухе;
- полноценно питаться;
- высыпаться;
- избегать стрессов;
- следить за балансом витаминов в организме.

Врач акушер-гинеколог, репродуктолог, консультант по грудному вскармливанию, окончила ЧГУ им. Ульянова со специализацией гинекология, маммология Подробнее »
* НА ЗАКАЗ: Купальник из двух частей в стиле ретро For Love & Lace
Главная > Продукты > * НА ЗАКАЗ: Купальник из двух частей в стиле ретро For Love & Lace
Верх
Белое кружево
Низ
Белое кружево
Цвет подкладки телесного цвета
Светло-коричневыйТемно-коричневый
Размер груди (самая полная часть)
Под грудью
Талия (дюймы)
Бедра (дюймы)
Окружность верхней части бедра
Рост
От промежности до пупка
Количество
Продано
НА ЗАКАЗ
Это модель на заказ. Каждое изделие изготавливается по индивидуальному размеру и цвету/печати каждого клиента. Поэтому возврат или обмен, к сожалению, не принимаются ни при каких условиях. Обязательно следуйте нашим 9 0003 инструкция по измерению и задать любой вопрос перед размещением заказа.
Каждое изделие изготавливается по индивидуальному размеру и цвету/печати каждого клиента. Поэтому возврат или обмен, к сожалению, не принимаются ни при каких условиях. Обязательно следуйте нашим 9 0003 инструкция по измерению и задать любой вопрос перед размещением заказа.
ОПИСАНИЕ
Красивые плавки в стиле ретро с завышенной талией из сексуальной кружевной вышивки из белой и телесной ткани с треугольным верхом в тон. Этот купальный костюм полностью на подкладке, изготовлен из ткани, растягивающейся в четырех направлениях, и может быть изготовлен с подкладкой телесного цвета двух разных цветов.
ТКАНЬ
Эластичность в 4 стороны, полностью на подкладке.
РАЗМЕР
Этот стиль изготавливается на заказ, и каждое изделие создано специально для вас. Пожалуйста, введите ваши точные размеры, чтобы мы могли убедиться, что купальник подходит правильно.
ДОСТАВКА
Пожалуйста, подождите 2-3 недели для доставки вашего индивидуального заказа в зависимости от текущего объема заказов. Если вы разместили заказ со стилями «в наличии», ваш заказ будет отправлен, когда заказ будет готов, чтобы избежать дополнительных затрат на доставку. Ваша посылка будет отправлена Priority Mail с номером отслеживания. Тарифы на доставку могут отличаться за пределами США.
Если вы разместили заказ со стилями «в наличии», ваш заказ будет отправлен, когда заказ будет готов, чтобы избежать дополнительных затрат на доставку. Ваша посылка будет отправлена Priority Mail с номером отслеживания. Тарифы на доставку могут отличаться за пределами США.
РАЗМЕР
Этот стиль изготавливается на заказ, и каждое изделие создано специально для вас. Пожалуйста, введите ваши точные размеры, чтобы мы могли убедиться, что купальник подходит правильно.
ДОСТАВКА
Пожалуйста, подождите 2-3 недели для доставки вашего индивидуального заказа в зависимости от текущего объема заказов. Если вы разместили заказ со стилями «в наличии», ваш заказ будет отправлен, когда заказ будет готов, чтобы избежать дополнительных затрат на доставку. Ваша посылка будет отправлена Priority Mail с номером отслеживания. Тарифы на доставку могут отличаться за пределами США.
ПРАВИЛА ВОЗВРАТА/ВОЗВРАТА/ОБМЕНА
Поскольку наши купальники изготавливаются на заказ по размеру и цвету каждого клиента, возврат или обмен, к сожалению, не принимаются ни при каких условиях. Мы не несем ответственности за неправильные измерения и, к сожалению, не сможем произвести обмен или возврат средств. Пожалуйста, обязательно следуйте нашим инструкциям по измерению и задавайте любые вопросы перед размещением заказа.
Мы не несем ответственности за неправильные измерения и, к сожалению, не сможем произвести обмен или возврат средств. Пожалуйста, обязательно следуйте нашим инструкциям по измерению и задавайте любые вопросы перед размещением заказа.
ИНСТРУКЦИИ ПО УХОДУ
Ручная стирка в холодной воде и развешивание для сушки.
Две маннозилтрансферазы KTR ответственны за биосинтез маннанов клеточной стенки и контроль поляризованного роста у Aspergillus fumigatus
1. Goto M. 2007. O-гликозилирование белков у грибов: разнообразные структуры и множественные функции. Биоски Биотехнолог Биохим 71:1415–1427. doi: 10.1271/bbb.70080. [PubMed] [CrossRef] [Google Scholar]
2. Дешпанде Н., Уилкинс М.Р., Пакер Н., Невалайнен Х. 2008. Пути гликозилирования белков у мицелиальных грибов. гликобиология 18: 626–637. дои: 10.1093/гликоб/cwn044. [PubMed] [CrossRef] [Google Scholar]
3. Mouyna I, Kniemeyer O, Jank T, Loussert C, Mellado E, Aimanianda V, Beauvais A, Wartenberg D, Sarfati J, Bayry J, Prévost M-C, Brakhage AA, Страль С. , Уэрре М., Латже Ж-П.
2010.
Члены семейства протеинов O-маннозилтрансфераз Aspergillus fumigatus по-разному влияют на рост, морфогенез и жизнеспособность. Мол Микробиол
76:1205–1221. doi: 10.1111/j.1365-2958.2010.07164.x. [PubMed] [CrossRef] [Академия Google]
, Уэрре М., Латже Ж-П.
2010.
Члены семейства протеинов O-маннозилтрансфераз Aspergillus fumigatus по-разному влияют на рост, морфогенез и жизнеспособность. Мол Микробиол
76:1205–1221. doi: 10.1111/j.1365-2958.2010.07164.x. [PubMed] [CrossRef] [Академия Google]
4. Вагенер Дж., Эхтенахер Б., Роде М., Коц А., Краппманн С., Хеземанн Дж., Эбель Ф. 2008. Предполагаемая α-1,2-маннозилтрансфераза AfMnt1 условно-патогенного грибкового патогена Aspergillus fumigatus необходима для стабильности клеточной стенки и полной вирулентности. эукариотическая клетка 7: 1661–1673. doi: 10.1128/EC.00221-08. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
5. Hall RA, Bates S, Lenardon MD, MacCallum DM, Wagener J, Lowman DW, Kruppa MD, Williams DL, Odds FC, Brown AJP, Gow НАР.
2013.
Семейство маннозилтрансфераз Mnn2 модулирует длину маннопротеиновых фибрилл, иммунное распознавание и вирулентность Кандида альбиканс . PLoS Патог
9:e1003276. doi: 10.1371/journal. ppat.1003276. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ppat.1003276. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
6. Goto M, Harada Y, Oka T, Matsumoto S, Takegawa K, Furukawa K. 2009. Белковые O-маннозилтрансферазы B и C поддерживают развитие и дифференцировку гиф у Aspergillus nidulans . эукариотическая клетка 8:1465–1474. doi: 10.1128/EC.00371-08. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Fontaine T, Simenel C, Dubreucq G, Adam O, Delepierre M, Lemoine J, Vorgias CE, Diaquin M, Latgé JP. 2000. Молекулярная организация щелоченерастворимой фракции Клеточная стенка Aspergillus fumigatus . J Биол Хим 275:27594–27607. doi: 10.1074/jbc.M909975199. [PubMed] [CrossRef] [Google Scholar]
8. Ламот Ф. 2016. Тестирование галактоманнана и 1,3-β-d-глюкана для диагностики инвазивного аспергиллеза. J Fungi (Базель) 2:22. doi: 10.3390/jof2030022. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Chai LYA, Vonk AG, Kullberg BJ, Verweij PE, Verschueren I, van der Meer JWM, Joosten LAB, Latgé J-P, Netea MG. 2011. Компоненты клеточной стенки Aspergillus fumigatus по-разному модулируют ответы TLR2 и TLR4 хозяина. микробы заражают
13:151–159. doi: 10.1016/j.micinf.2010.10.005. [PubMed] [CrossRef] [Google Scholar]
2011. Компоненты клеточной стенки Aspergillus fumigatus по-разному модулируют ответы TLR2 и TLR4 хозяина. микробы заражают
13:151–159. doi: 10.1016/j.micinf.2010.10.005. [PubMed] [CrossRef] [Google Scholar]
10. Bozza S, Clavaud C, Giovannini G, Fontaine T, Beauvais A, Sarfati J, D’Angelo C, Perruccio K, Bonifazi P, Zagarella S, Moretti S, Bistoni Ф, Латге Ж-П, Романи Л. 2009. Иммунное зондирование белков, гликолипидов и полисахаридов Aspergillus fumigatus и влияние на иммунитет Th и вакцинацию. Дж Иммунол 183:2407–2414. дои: 10.4049/jиммунол.0
1. [PubMed] [CrossRef] [Google Scholar]
11. Манро С. 2001. Что могут рассказать нам дрожжи о N-связанном гликозилировании в аппарате Гольджи. FEBS Lett 498: 223–227. [PubMed] [Google Scholar]
12. Latgé JP, Kobayashi H, Debeaupuis JP, Diaquin M, Sarfati J, Wieruszeski JM, Parra E, Bouchara JP, Fournet B.
1994.
Химическая и иммунологическая характеристика внеклеточного галактоманнана Aspergillus fumigatus . Заразить иммунитет
62:5424–5433. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Заразить иммунитет
62:5424–5433. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Генри С., Фонтейн Т., Хеддерготт С., Робине П., Айманианда В., Бо Р., Бове А., Муйна И., Прево М.С., Феккар А., Чжао И., Перлин Д., Латге Ж.-П. 2016. Биосинтез маннана клеточной стенки в конидии и мицелии Aspergillus fumigatus . Клеточная микробиология 18:1881–1891. дои: 10.1111/см.12665. [PubMed] [CrossRef] [Google Scholar]
14. Lambou K, Perkhofer S, Fontaine T, Latge J-P. 2010. Сравнительный функциональный анализ семейств маннозилтрансфераз OCh2 в Aspergillus fumigatus и Saccharomyces cerevisiae . Дрожжи 27:625–636. doi: 10.1002/yea.1798. [PubMed] [CrossRef] [Google Scholar]
15. Lussier M, Sdicu AM, Bussereau F, Jacquet M, Bussey H. 1997. Маннозилтрансферазы Ktr1p, Ktr3p и Kre2p/Mnt1p участвуют в выработке O- и N-связанных углеводных цепей дрожжей. J Биол Хим 272:15527–15531.] [PubMed] [Google Scholar]
16. Mouyna I, Aimanianda V, Hartl L, Prevost M-C, Sismeiro O, Dillies MA, Jagla B, Legendre R, Coppee JY, Latgé J-P. 2016.
β-(1,3)-глюканазы семейства Gh26 и GH81 в Aspergillus fumigatus необходимы для морфогенеза конидиальной клеточной стенки. Клеточная микробиология
18:1285–1293. дои: 10.1111/см.12630. [PubMed] [CrossRef] [Google Scholar]
2016.
β-(1,3)-глюканазы семейства Gh26 и GH81 в Aspergillus fumigatus необходимы для морфогенеза конидиальной клеточной стенки. Клеточная микробиология
18:1285–1293. дои: 10.1111/см.12630. [PubMed] [CrossRef] [Google Scholar]
17. Hartmann T, Dümig M, Jaber BM, Szewczyk E, Olbermann P, Morschhäuser J, Krappmann S. 2010. Валидация маркера самоиссечения в патогене человека Aspergillus fumigatus с использованием системы рекомбинации бета-рек/шесть сайтов. Appl Environ Microbiol 76:6313–6317. doi: 10.1128/AEM.00882-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Морель В., Бернар М., Дебопюи Ж.-П., Буитраго М., Табуре М., Латже Ж.-П. 2005. Галактоманнопротеины Aspergillus fumigatus . эукариотическая клетка 4: 1308–1316. doi: 10.1128/EC.4.7.1308-1316.2005. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
19. Krüger AT, Engel J, Buettner FFR, Routier FH.
2016. Aspergillus fumigatus Cap59-подобный белок А участвует в α1,3-маннозилировании GPI-якорей.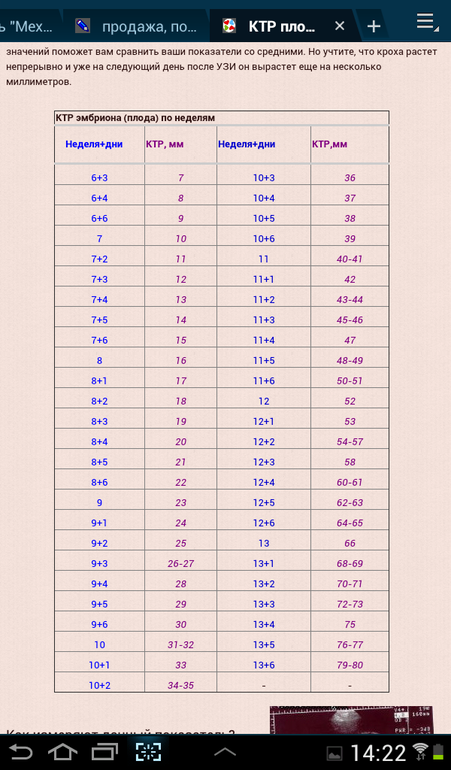 гликобиология
26:30–38. doi: 10.1093/гликоб/cwv078. [PubMed] [CrossRef] [Академия Google]
гликобиология
26:30–38. doi: 10.1093/гликоб/cwv078. [PubMed] [CrossRef] [Академия Google]
20. Ризон А.Дж., Делл А., Ромеро П.А., Херскович А. 1991. Специфичность маннозилтрансферазы, которая инициирует образование внешней цепи у Saccharomyces cerevisiae . гликобиология 1: 387–391. [PubMed] [Google Scholar]
21. Родионов Д., Ромеро П.А., Бергуис А.М., Херскович А. 2009. Экспрессия и очистка рекомбинантного M-Pol I из Saccharomyces cerevisiae с активностью α-1,6 маннозилполимеразы. Protein Expr Purif 66:1–6. doi: 10.1016/j.pep.2009.02.013. [PubMed] [CrossRef] [Академия Google]
22. Юнгманн Дж., Манро С. 1998. Мультибелковые комплексы в цис-Гольджи Saccharomyces cerevisiae с активностью альфа-1,6-маннозилтрансферазы. ЭМБО J 17:423–434. doi: 10.1093/emboj/17.2.423. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Jungmann J, Rayner JC, Munro S.
1999.
Белок Saccharomyces cerevisiae Mnn10p/Bed1p является субъединицей маннозилтрансферазного комплекса Гольджи. J Биол Хим
274: 6579–6585. [PubMed] [Академия Google]
J Биол Хим
274: 6579–6585. [PubMed] [Академия Google]
24. Штольц Дж., Манро С. 2002. Компоненты маннозилтрансферазного комплекса M-Pol I Saccharomyces cerevisiae выполняют различные функции в синтезе маннана. J Биол Хим 277:44801–44808. doi: 10.1074/jbc.M208023200. [PubMed] [CrossRef] [Google Scholar]
25. Wang JJ, Qiu L, Cai Q, Ying S-H, Feng M-G. 2014. Три α-1,2-маннозилтрансферазы по-разному способствуют конидиации, целостности клеточной стенки, устойчивости к множественным стрессам и вирулентности Beauveria bassiana . Грибковый Генет Биол 70:1–10. doi: 10.1016/j.fgb.2014.06.010. [PubMed] [CrossRef] [Google Scholar]
26. Muszkieta L, Aimanianda V, Mellado E, Gribaldo S, Alcàzar-Fuoli L, Szewczyk E, Prevost MC, Latgé J-P.
2014.
Расшифровка роли семейств хитинсинтаз 1 и 2 в росте in vivo и in vitro Aspergillus fumigatus путем множественной делеции гена. Клеточная микробиология
16: 1784–1805. дои: 10.1111/см. 12326. [PubMed] [CrossRef] [Академия Google]
12326. [PubMed] [CrossRef] [Академия Google]
27. Муйна И., Фонтейн Т., Вай М., Моно М., Фонзи В.А., Дьякин М., Пополо Л., Хартланд Р.П., Латге Д.П. 2000. Гликозилфосфатидилинозитол-заякоренные глюканозилтрансферазы играют активную роль в биосинтезе клеточной стенки грибов. J Биол Хим 275:14882–14889. [PubMed] [Google Scholar]
28. Gastebois A, Fontaine T, Latgé J-P, Mouyna I. 2010. β(1-3)Glucanosyltransferase Gel4p необходим для Aspergillus fumigatus . эукариотическая клетка 9: 1294–1298. doi: 10.1128/EC.00107-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Такэсита Н. 2016. Координированный процесс поляризованного роста у мицелиальных грибов. Биоски Биотехнолог Биохим 80: 1693–1699. дои: 10.1080/09168451.2016.1179092. [PubMed] [CrossRef] [Google Scholar]
30. Riquelme M, Aguirre J, Bartnicki-García S, Braus GH, Feldbrügge M, Fleig U, Hansberg W, Herrera-Estrella A, Kämper J, Kück U, Mouriño- Перес Р.Р., Такешита Н., Фишер Р.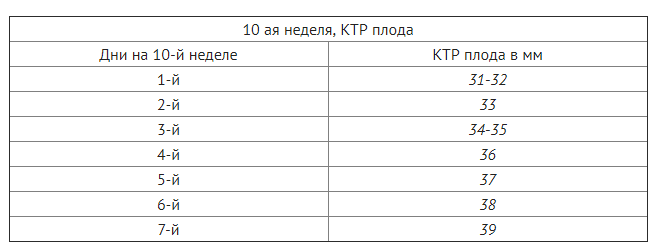 2018.
Морфогенез грибов, от поляризованного роста гиф до сложных структур размножения и заражения. Микробиол Мол Биол Рев
82:e00068-17. doi: 10.1128/MMBR.00068-17. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2018.
Морфогенез грибов, от поляризованного роста гиф до сложных структур размножения и заражения. Микробиол Мол Биол Рев
82:e00068-17. doi: 10.1128/MMBR.00068-17. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Харрис С.Д., Момани М. 2004. Полярность мицелиальных грибов: выход за рамки парадигмы дрожжей. Грибковый Генет Биол 41:391–400. doi: 10.1016/j.fgb.2003.11.007. [PubMed] [CrossRef] [Google Scholar]
32. Oda K, Bignell E, Kang SE, Momany M. 2017. Уровни транскриптов модуля Aspergillus fumigatus Cdc42, поляризомы и генов септина мало изменяются от состояния покоя до установления полярности. Мед Микол 55:445–452. дои: 10.1093/mmy/myw085. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Дихтл К., Самантарай С., Айманианда В., Чжу З., Прево М.-К., Латге Дж.-П., Эбель Ф., Вагенер Дж.
2015. Aspergillus fumigatus лишенный клеточной стенки β-1,3-глюкан является жизнеспособным, массово выделяет галактоманнан и уничтожается ингибиторами образования перегородки. Мол Микробиол
95:458–471. дои: 10.1111/mmi.12877. [PubMed] [CrossRef] [Google Scholar]
Мол Микробиол
95:458–471. дои: 10.1111/mmi.12877. [PubMed] [CrossRef] [Google Scholar]
34. Henry C, Latgé J-P, Beauvais A. 2012. α1,3-глюканы незаменимы в Aspergillus fumigatus. эукариотическая клетка 11:26–29. doi: 10.1128/EC.05270-11. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Gravelat FN, Beauvais A, Liu H, Lee MJ, Snarr BD, Chen D, Xu W, Kravtsov I, Hoareau CMQ, Vanier G, Urb M, Campoli P, Al Abdallah Q, Lehoux M, Chabot JC, Ouimet MC, Baptista SD, Fritz JH, Nierman WC, Latgé JP, Mitchell AP, Filler SG, Fontaine T, Sheppard DC. 2013. Галактозаминогалактан Aspergillus опосредует адгезию к компонентам хозяина и скрывает β-глюкан гиф от иммунной системы. PLoS Патог 9:e1003575. doi: 10.1371/journal.ppat.1003575. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Самар Д., Килер Дж. Б., Клутц Дж. С.
2015.
Идентификация и делеция Tft1, предполагаемой гликозилтрансферазы, необходимой для синтеза β-1,3;1,4-глюкана клеточной стенки у Aspergillus fumigatus . PLoS Один
10:e0117336. doi: 10.1371/journal.pone.0117336. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
PLoS Один
10:e0117336. doi: 10.1371/journal.pone.0117336. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Komachi Y, Hatakeyama S, Motomatsu H, Futagami T, Kizjakina K, Sobrado P, Ekino K, Takegawa K, Goto M, Nomura Y, Oka Т. 2013. gfsA кодирует новую галактофуранозилтрансферазу, участвующую в биосинтезе галактофуранозного антигена О-гликана в Aspergillus nidulans и A. fumigatus . Мол Микробиол 90:1054–1073. doi: 10.1111/mmi.12416. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Katafuchi Y, Li Q, Tanaka Y, Shinozuka S, Kawamitsu Y, Izumi M, Ekino K, Mizuki K, Takegawa K, Shibata N, Goto М, Номура Ю, Охта К, Ока Т. 2017. GfsA представляет собой β1,5-галактофуранозилтрансферазу, участвующую в биосинтезе боковой цепи галактофурана галактоманнана грибкового типа в Aspergillus fumigatus 9.0100 . гликобиология 27: 568–581. doi: 10.1093/гликоб/cwx028. [PubMed] [CrossRef] [Google Scholar]
39. Lamarre C, Beau R, Balloy V, Fontaine T, Hoi JWS, Guadagnini S, Berkova N, Chignard M, Beauvais A, Latgé J-P. 2009.
Галактофураноза ослабляет клеточную адгезию Aspergillus fumigatus . Клеточная микробиология
11: 1612–1623. doi: 10.1111/j.1462-5822.2009.01352.x. [PubMed] [CrossRef] [Google Scholar]
2009.
Галактофураноза ослабляет клеточную адгезию Aspergillus fumigatus . Клеточная микробиология
11: 1612–1623. doi: 10.1111/j.1462-5822.2009.01352.x. [PubMed] [CrossRef] [Google Scholar]
40. Engel J, Schmalhorst PS, Routier FH. 2012. Для биосинтеза полисахарида галактоманнана клеточной стенки грибов требуется внутрипросветная ГДФ-манноза. J Биол Хим 287:44418–44424. дои: 10.1074/jbc.M112.398321. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Lussier M, Sdicu AM, Bussey H. 1999. Семейства маннозилтрансфераз KTR и MNN1 из Saccharomyces cerevisiae . Биохим Биофиз Акта 1426: 323–334. [PubMed] [Google Scholar]
42. Rayner JC, Munro S. 1998. Идентификация маннозилтрансфераз MNN2 и MNN5, необходимых для формирования и удлинения маннозных ветвей маннанов внешней цепи Saccharomyces cerevisiae . J Биол Хим 273:26836–26843. [PubMed] [Академия Google]
43. Сфихи-Луалия Г., Юрто Т., Фабр Э., Фраден С., Мее А., Пурсело М. , Маес Э., Букарт Дж., Малле Ж-М, Пулен Д., Дельплас Ф., Герардель Ю.
2016. Candida albicans β-1,2-маннозилтрансфераза Bmt3 вызывает удлинение фосфопептидоманнана клеточной стенки. гликобиология
26:203–214. doi: 10.1093/гликоб/cwv094. [PubMed] [CrossRef] [Google Scholar]
, Маес Э., Букарт Дж., Малле Ж-М, Пулен Д., Дельплас Ф., Герардель Ю.
2016. Candida albicans β-1,2-маннозилтрансфераза Bmt3 вызывает удлинение фосфопептидоманнана клеточной стенки. гликобиология
26:203–214. doi: 10.1093/гликоб/cwv094. [PubMed] [CrossRef] [Google Scholar]
44. da Silva Ferreira ME, Kress MR, Savoldi M, Goldman MH, Härtl A, Heinekamp T, Brakhage AA, Goldman GH. 2006. Мутант akuBKU80, лишенный негомологичного соединения концов, является мощным инструментом для анализа патогенности Aspergillus fumigatus. эукариотическая клетка 5:207–211. doi: 10.1128/EC.5.1.207-211.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Коув Диджей. 1966 год. Индукция и репрессия нитратредуктазы у гриба Aspergillus nidulans . Биохим Биофиз Акта 113:51–56. [PubMed] [Google Scholar]
46. Girardin H, Latgé JP, Srikantha T, Morrow B, Soll DR.
1993.
Разработка ДНК-зондов для снятия отпечатков пальцев Aspergillus fumigatus . Джей Клин Микробиол
31: 1547–1554. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Джей Клин Микробиол
31: 1547–1554. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Сезонов Г., Жозеле-Пти Д., Д’Ари Р. 2007. Кишечная палочка физиология в бульоне Луриа-Бертани. J Бактериол 189:8746–8749. дои: 10.1128/JB.01368-07. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Clavaud C, Beauvais A, Barbin L, Munier-Lehmann H, Latgé J-P. 2012. Состав питательной среды влияет на метаболизм β-1,3-глюканов Aspergillus fumigatus и противогрибковую активность ингибиторов синтеза β-1,3-глюканов. Противомикробные агенты Chemother 56:3428–3431. doi: 10.1128/AAC.05661-11. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Дюбуа М., Жиль К.А., Гамильтон Дж.К., Реберс П.А., Смит Ф. 1956. Колориметрический метод определения сахаров и родственных им веществ. Анальная химия 28:350–356. doi: 10.1021/ac60111a017. [CrossRef] [Google Scholar]
50. Stalhberger T, Simenel C, Clavaud C, Eijsink VGH, Jourdain R, Delepierre M, Latgé J-P, Breton L, Fontaine T. 2014.
Химическая организация полисахаридного ядра клеточной стенки Malassezia Restricta . J Биол Хим
289:12647–12656. doi: 10.1074/jbc.M113.547034. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2014.
Химическая организация полисахаридного ядра клеточной стенки Malassezia Restricta . J Биол Хим
289:12647–12656. doi: 10.1074/jbc.M113.547034. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Морель В., Фаид В., Чират Ф., Михальски Дж.-К. 2009. Анализ N- и O-связанных гликанов из гликопротеинов с помощью масс-спектрометрии MALDI-TOF. Методы Мол Биол 534:5–21. doi: 10.1007/978-1-59745-022-5_1. [PubMed] [CrossRef] [Google Scholar]
52. Striebeck A, Robinson DA, Schüttelkopf AW, van Aalten DMF. 2013. Mnn9 дрожжей является одновременно праймирующей гликозилтрансферазой и аллостерическим активатором биосинтеза маннана. Открытая биология 3:130022. doi: 10.1098/rsob.130022. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Хилл К., Бун С., Гебл М., Пучча Р., Сиду А.М., Бусси Х.
1992.
Yeast Kre2 определяет новое семейство генов, кодирующих вероятные секреторные белки, и требуется для правильного N-гликозилирования белков. Генетика
130: 273–283.