пошаговая инструкция в 2022 году
Работник принес больничный лист, но болел не сам, а ухаживал за ребенком. Есть особенности при назначении пособия по временной нетрудоспособности
Главные правила бухгалтера
- Назначение и выплата пособия по уходу за ребенком производятся полностью (с 1-го дня нетрудоспособности) за счет средств ФСС (ч. 3 ст. 3 Федерального закона № 255-ФЗ). Работодатель не обязан оплачивать за счет собственных средств первые три дня нетрудоспособности.
- Размер и порядок назначения пособия по временной нетрудоспособности зависят от возраста ребенка, вида заболевания, места лечения ребенка (ч. 5 ст. 6 Закона от 29.12.2006 № 255-ФЗ).
- Дни больничного по уходу за ребенком, приходящиеся на время ежегодного отпуска, не оплачиваются. Пособие по больничному полагается со дня, когда работник должен был выйти на работу (ст. 9 Закона № 255-ФЗ). Оплачивают только те дни временной нетрудоспособности, которые приходятся на дни после окончания ежегодного отпуска.


Назначение пособия в 2022 году. Алгоритм взаимодействия работодателя с ФСС
С 1 января 2022 года порядок передачи работодателем сведений в ФСС РФ для назначения и выплаты пособия по временной нетрудоспособности регламентирует Постановление Правительства РФ от 23.11.2021 № 2010.
- Необходимо посмотреть информацию в сервисе электронного взаимодействия с ФСС — «Социальный электронный документооборот» (СЭДО). Этот сервис интегрирован в бухгалтерские программы.
- Если информация о закрытии больничного листа есть, то в течение 3 рабочих дней по запросу работодателю необходимо передать сведения для выплаты пособия в ФСС РФ. Перечень сведений, предоставляемых в ФСС РФ, поименован в п. 22 Постановления Правительства РФ от 23.11.2021 № 2010. Работодатель предоставляет Фонду только те сведения, которые будут указаны в запросе. При наличии у ФСС РФ необходимых ему для назначения пособия сведений у работодателя они не запрашиваются.
- В течение 5 рабочих дней при необходимости специалист ФСС РФ может запросить у работодателя дополнительные сведения для назначения пособия, Например информацию о районном коэффициенте.
- Если поступит запрос ФСС о предоставлении дополнительных сведений, работодателю необходимо:
- в течение 3 рабочих дней подтвердить, что он получил запрос;
- в течение 5 рабочих дней предоставить дополнительные сведения.

С 1 января 2022 года срок выплаты территориальным органом ФСС РФ суммы пособия по временной нетрудоспособности составляет 10 рабочих дней со дня получения им сведений, которые необходимы для назначения и выплаты пособия.
В конце статьи есть шпаргалка
Количество дней нетрудоспособности, которые можно оплатить
Количество дней нетрудоспособности, которые будут оплачены работнику, зависит:
- от возраста ребенка;
- от вида заболевания ребенка;
- от места лечения ребенка: амбулаторно или в стационаре.

В ст. 6 Федерального закона от 29.12.2006 № 255-ФЗ указаны условия и продолжительность выплаты пособия по временной нетрудоспособности, в том числе в случае, если работник осуществлял уход за ребенком. Рассмотрим их в таблице.
| Возраст ребенка | Количество оплачиваемых дней | Ограничения | Код причины нетрудоспособности | Особенности |
|---|---|---|---|---|
| До 7 лет | Весь период нетрудоспособности независимо от места лечения (амбулаторно или в стационаре) | Не более 90 календарных дней в календарном году по всем случаям ухода за этим ребенком | «12» | Заболевание включено в перечень, утв. Приказом Минздравсоцразвития РФ от 20.02.2008 № 84н |
| Не более 60 календарных дней в календарном году по всем случаям ухода за этим ребенком | «09» | |||
| От 7 до 15 лет | За период до 15 календарных дней по каждому случаю лечения ребенка амбулаторно или в стационаре | Не более 45 календарных дней в календарном году по всем случаям ухода за ребенком | ||
| 15 лет и старше | Не более 7 календарных дней по каждому случаю заболевания при лечении амбулаторно | Не более 30 календарных дней в календарном году по всем случаям ухода за этим ребенком | ||
| В возрасте до 18 лет в случае ухода за больным ребенком-инвалидом | Весь период нетрудоспособности независимо от места лечения (амбулаторно или в стационаре) | Не более 120 календарных дней в календарном году по всем случаям ухода за этим ребенком | «13» | |
| В возрасте до 18 лет при его болезни, связанной с поствакцинальным осложнением, при злокачественных новообразованиях | Весь период нетрудоспособности независимо от места лечения (амбулаторно или в стационаре) | «14» | ||
| В возрасте до 18 лет, если ребенок является ВИЧ-инфицированным | Весь период нетрудоспособности (только в стационаре) | «15» |
Внимание: законодательство РФ не регулирует вопрос, кто должен вести подсчет дней временной нетрудоспособности по уходу за больным ребенком за соответствующий календарный год.
Мнение ФСС РФ: учет количества дней, за которые работнику должно быть выплачено пособие по временной нетрудоспособности в связи с необходимостью ухода за больным ребенком, входит в обязанности работодателя (Письмо ФСС РФ от 19.12.2014 № 17-03-14/06-18772).
Онлайн-курс в Контур.Школе
Расчеты с персоналом по оплате труда. 146 ак. часов. Удостоверение
Посмотреть программуСовет работодателю: как учитывать и контролировать количество использованных дней по уходу за больным ребенком:
- завести специальный журнал регистрации в удобной для него форме; или
- вести учет таких дней в таблице (например, в Exсel).
Размер пособия по временной нетрудоспособности
С 01.09.2021 года размер пособия по временной нетрудоспособности зависит не только от страхового стажа работника, но и от возраста ребенка. Порядок определения размера пособия приведен в ст. 7 Федерального Закона от 29.12.2006 № 255-ФЗ.
- Ситуация 1. Возраст больного ребенка до 8 лет. Весь период нетрудоспособности оплачивается работнику исходя из 100% среднего заработка. Стаж работника здесь значения не имеет.
- при лечении ребенка амбулаторно первые 10 календарных дней нетрудоспособности оплачиваются в размере, определяемом в зависимости от продолжительности страхового стажа работника, а последующие дни — в размере 50% от среднего заработка;
- при лечении ребенка в стационаре пособие выплачивается в размере, определяемом в зависимости от продолжительности страхового стажа работника.
При лечении амбулаторно ребенка в возрасте 15 лет и старше пособие выплачивается не более чем за 7 календарных дней в размере, зависящем от продолжительности страхового стажа.
Размер среднего заработка в зависимости от страхового стажа работника (ч.
- страховой стаж 8 и более лет — выплачивается 100% среднего заработка;
- от 5 до 8 лет — 80% среднего заработка;
- до 5 лет — 60% среднего заработка.
Пример
В ООО «Прогресс» работает Ложкина Вера. С 18 по 31 августа 2022 года (14 календарных дней) она была на больничном по уходу за ребенком, которому 9 лет.
В 2022 году Ложкина уже была на больничном по уходу за этим ребенком:
- с 10 по 16 марта — 7 дней;
- с 4 по 20 апреля — 17 дней;
- с 14 по 29 июня — 16 дней.
- Вере Ложкиной оплатят первые 5 дней нетрудоспособности (с 18 по 22 августа) в зависимости от ее страхового стажа, а оставшиеся 9 дней (с 23 по 31 августа) — нет.
Шпаргалка
В шпаргалке собрана полезная информация из статьи:
-
Пример расчета больничного в 2022 году 645.
9 КБ
 9 КБ
9 КБ
Как оплачивается больничный лист по уходу за больным ребенком
С 1 сентября 2021 года пособие по временной нетрудоспособности
при необходимости осуществления ухода за больным ребенком в возрасте
до 8 лет будет выплачиваться в размере 100% среднего заработка независимо
от страхового стажа
Для получения пособия по временной нетрудоспособности в связи
с необходимостью ухода за больным ребенком работник должен предоставить работодателю больничный лист. Пособие назначает и выплачивает территориальный орган ФСС РФ. Не всегда оплачивается весь период освобождения от работы. Максимальное количество дней, которые могут быть оплачены, зависит от возраста ребенка и вида его заболевания. Также имеет значение, где лечили ребенка: дома или в стационаре.
Пособие по временной нетрудоспособности в связи с уходом за ребенком назначает и выплачивает территориальный орган ФСС РФ за счет средств бюджета фонда. Для назначения пособия организация представляет в ФСС РФ документы (сведения), в том числе сведения о застрахованном лице, а также
Для назначения пособия организация представляет в ФСС РФ документы (сведения), в том числе сведения о застрахованном лице, а также
(в зависимости от способа подачи) опись представленных документов (сведений) или реестр сведений (п. п. 2, 3, 10 Положения об особенностях назначения
и выплаты в 2021 г. страхового обеспечения по ВНиМ).
Выходные дни в больничном листе оплачиваются наравне с рабочими, если они включены в период нетрудоспособности. В ч. 1 ст. 9 Закона N 255-ФЗ перечислены периоды, когда пособие по временной нетрудоспособности
не назначается. Выходные и нерабочие праздничные дни в этот перечень не входят (Письмо ФСС РФ от 31.01.2017 N 02-09-14/22-03-848).
Максимальное количество оплачиваемых дней больничного по уходу
за ребенком зависит от возраста ребенка и вида его заболевания (ч. 5 ст. 6 Закона N 255-ФЗ).
Больничный лист по уходу за ребенком выдается и оплачивается матери, отцу, бабушке, другим родственникам, опекуну, попечителю (п. 40 Порядка выдачи
40 Порядка выдачи
и оформления листков нетрудоспособности). Степень родства не влияет
на порядок оплаты.
Если работник, находящийся в отпуске по уходу за ребенком, не работает,
то больничный лист по уходу за больным ребенком не оплачивается (п. 1 ч. 1
ст. 9 Закона N 255-ФЗ).
Если сумма пособия в расчете за полный календарный месяц окажется меньше МРОТ, пособие выплачивается в размере, исчисленном из МРОТ (ч. 6.1
ст. 14 Закона N 255-ФЗ).
При лечении в стационаре ребенка 15 лет и старше (как и в случае лечения
в стационаре другого члена семьи) больничный лист по уходу не оформляют
(п. 46 Порядка выдачи и оформления листков нетрудоспособности).
Можно ли не сдавать электронный больничный?
Главная → Статьи → Можно ли не сдавать электронный больничный?
Сотрудник получил электронный больничный, но все основное время был на работе, в табеле учета рабочего времени отмечено, что сотрудник работал полный рабочий день.
Может ли сотрудник не сдавать электронный больничный? Является ли это нарушением с какой-либо стороны?
Согласно ст. 183 ТК РФ при временной нетрудоспособности работодатель выплачивает работнику пособие по временной нетрудоспособности в соответствии с федеральными законами. Условия его выплаты устанавливаются Федеральным законом от 29.12.2006 № 255-ФЗ “Об обязательном социальном страховании на случай временной нетрудоспособности и в связи с материнством” (далее – Закон № 255-ФЗ).
В соответствии с ч. 8 ст. 6 Закона № 255-ФЗ пособие по временной нетрудоспособности выплачивается застрахованному лицу во всех случаях за календарные дни, приходящиеся на соответствующий период, за исключением календарных дней, приходящихся на периоды, указанные в ч. 1 ст. 9 Закона № 255-ФЗ. Следовательно, факт наступления у работника временной нетрудоспособности порождает у работодателя обязанность по выплате соответствующего пособия.
Основанием для назначения работодателем работнику пособия по временной нетрудоспособности является листок нетрудоспособности, выданный медицинской организацией по форме и в порядке (далее – Порядок), утвержденным соответственно приказами Минздравсоцразвития РФ от 26. 04.2011 № 347н и от 29.06.2011 № 624н (ч.ч. 1, 5 ст. 13 Закона № 255-ФЗ). При этом согласно ч. 3.2 ст. 59 Федерального закона от 21.11.2011 № 323-ФЗ “Об основах охраны здоровья граждан в Российской Федерации”, листок нетрудоспособности может выдаваться как в форме документа на бумажном носителе, так и (с письменного согласия пациента) в виде электронного документа, подписанного с использованием усиленной квалифицированной электронной подписи медицинским работником и медицинской организацией. Это возможно, если медицинская организация и страхователь являются участниками системы информационного взаимодействия по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа.
04.2011 № 347н и от 29.06.2011 № 624н (ч.ч. 1, 5 ст. 13 Закона № 255-ФЗ). При этом согласно ч. 3.2 ст. 59 Федерального закона от 21.11.2011 № 323-ФЗ “Об основах охраны здоровья граждан в Российской Федерации”, листок нетрудоспособности может выдаваться как в форме документа на бумажном носителе, так и (с письменного согласия пациента) в виде электронного документа, подписанного с использованием усиленной квалифицированной электронной подписи медицинским работником и медицинской организацией. Это возможно, если медицинская организация и страхователь являются участниками системы информационного взаимодействия по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа.
В письме ФСС РФ от 11.08.2017 № 02-09-11/22-05-13462 отмечается, что электронный листок нетрудоспособности (далее – ЭЛН) с 01.07.2017 имеет равную юридическую силу с листком нетрудоспособности на бумажном носителе.
На сегодняшний день порядок взаимодействия между медицинской организацией, застрахованным (работником), страховщиком (работодателем) и страхователем (ФСС) еще не утвержден. Минтрудом России подготовлен проект постановления Правительства РФ “Об утверждении Порядка информационного взаимодействия страховщика, страхователей, медицинских организаций и федеральных государственных учреждений медико-социальной экспертизы по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа”. Также разработан проект постановления Правительства РФ “Об утверждении Правил информационного взаимодействия страховщика, страхователей, медицинских организаций и федеральных государственных учреждений медико-социальной экспертизы по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа”.
Минтрудом России подготовлен проект постановления Правительства РФ “Об утверждении Порядка информационного взаимодействия страховщика, страхователей, медицинских организаций и федеральных государственных учреждений медико-социальной экспертизы по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа”. Также разработан проект постановления Правительства РФ “Об утверждении Правил информационного взаимодействия страховщика, страхователей, медицинских организаций и федеральных государственных учреждений медико-социальной экспертизы по обмену сведениями в целях формирования листка нетрудоспособности в форме электронного документа”.
На официальном сайте Фонда социального страхования РФ размещены Памятки для застрахованного лица, медицинской организации, страхователя по формированию ЭЛН.
1. Работник, обратившись в медицинское учреждение, сообщает свой СНИЛС и дает согласие на формирование электронного больничного.
2. Медицинская организация открывает ЭЛН и присваивает ему индивидуальный номер.
3. После выписки работник сообщает номер ЭЛН работодателю.
4. Работодатель при помощи выбранного метода информационного взаимодействия запрашивает у ФСС данные на работника по номеру его СНИЛС и ЭЛН и т.д.
Таким образом, листок нетрудоспособности в любой его форме является надлежащим документом для подтверждения факта временной нетрудоспособности, а также права застрахованного лица на получение пособия по временной нетрудоспособности. Однако в рассматриваемом случае работник продолжил работу и не предоставил работодателю информацию о номере ЭЛН, оформленного в медицинской организации, куда он обратился за медицинской помощью. Такие действия свидетельствуют о том, что работник добровольно не воспользовался правом на освобождение от работы в связи с нетрудоспособностью.
Если работник добровольно приступает к труду в период, когда он, по мнению врача, нетрудоспособен, у работодателя нет оснований не допускать его к работе, поскольку такое основание, как временная нетрудоспособность работника, не поименовано среди оснований отстранения от работы (ст. 76 ТК РФ), за исключением отстранения от работы в соответствии с медицинским заключением, согласно которому работнику противопоказано выполнение работы, обусловленной трудовым договором (что не относится к рассматриваемому случаю).
76 ТК РФ), за исключением отстранения от работы в соответствии с медицинским заключением, согласно которому работнику противопоказано выполнение работы, обусловленной трудовым договором (что не относится к рассматриваемому случаю).
Соблюдение работником режима в период его временной нетрудоспособности, предписанного ему врачом, трудовой обязанностью работника не является. Трудовое законодательство не устанавливает ни способы контроля со стороны работодателя за соблюдением режима при временной нетрудоспособности, ни право контролировать соблюдение работником такого режима. При этом на работодателе лежит обязанность учитывать отработанное работником время (часть четвертая ст. 91 ТК РФ). Кроме того, работодатель обязан выплачивать работникам заработную плату за отработанное ими время (ст.ст. 56, 129, 132, 135 ТК РФ).
Поскольку работник в дни освобождения от работы в связи с нетрудоспособностью исполнял свои трудовые обязанности и листок нетрудоспособности работодателю не представил, то работодатель правомерно учел в табеле учета рабочего времени это время как отработанное и поэтому за указанный период должен выплатить заработную плату. Последующее сообщение работодателю номера ЭЛН, по нашему мнению, не должно повлечь за собой каких-либо юридических последствий и не потребует от работодателя совершать дальнейшие действия по начислению работнику пособия по временной нетрудоспособности (смотрите, например, апелляционное определение СК по гражданским делам Тверского областного суда от 08.04.2016 по делу № 33-854/2016). Пособие по временной нетрудоспособности призвано компенсировать гражданам утраченный заработок в связи с наступлением временной нетрудоспособности (п. 1 ч. 1 ст. 1.2, п. 1 ч. 2 ст. 1.3, п. 1 ч. 1 ст. 1.4 Закона № 255-ФЗ). Поэтому выплата пособия за период, в течение которого заработок не утрачен, невозможна и противоречит законодательству (смотрите, например, апелляционное определение СК по гражданским делам Хабаровского краевого суда от 30.03.2016 по делу № 33-2279/2016).
Последующее сообщение работодателю номера ЭЛН, по нашему мнению, не должно повлечь за собой каких-либо юридических последствий и не потребует от работодателя совершать дальнейшие действия по начислению работнику пособия по временной нетрудоспособности (смотрите, например, апелляционное определение СК по гражданским делам Тверского областного суда от 08.04.2016 по делу № 33-854/2016). Пособие по временной нетрудоспособности призвано компенсировать гражданам утраченный заработок в связи с наступлением временной нетрудоспособности (п. 1 ч. 1 ст. 1.2, п. 1 ч. 2 ст. 1.3, п. 1 ч. 1 ст. 1.4 Закона № 255-ФЗ). Поэтому выплата пособия за период, в течение которого заработок не утрачен, невозможна и противоречит законодательству (смотрите, например, апелляционное определение СК по гражданским делам Хабаровского краевого суда от 30.03.2016 по делу № 33-2279/2016).
Таким образом, нарушений законодательства со стороны работника, не сообщившего работодателю сведений об оформленном листке нетрудоспособности, и отработавшего предусмотренные его режимом работы рабочие часы, и работодателя, зафиксировавшего отработанное работником время, в рассматриваемом случае нет.
Ответ подготовил: Жгулева Ольга, эксперт службы Правового консалтинга ГАРАНТ
Контроль качества ответа: Комарова Виктория, рецензент службы Правового консалтинга ГАРАНТ
Свежие новости цифровой экономики на нашем канале в Телеграм
| Нужна электронная подпись? Достаточно оставить заявку. Мы поможем выбрать нужный в вашем случае тип сертификата электронной подписи, расскажем как его применить и предоставим другие дополнительные услуги. Оставить заявку >> |
Гериатры призывают не нырять в прорубь без подготовки
Врачи Российского геронтологического научно-клинического центра РНИМУ им. Н.И. Пирогова сообщили, что ныряние в прорубь в пожилом возрасте может привести к ухудшению когнитивных функций, остановке сердца и смерти.
Заведующий отделением гериатрической реабилитации Российского геронтологического научно-клинического центра РНИМУ им. Н.И. Пирогова, врач-физиотерапевт, врач ЛФК Александр Мелия предупредил, что в пожилом возрасте ныряние в прорубь без соответствующей подготовки может привести к остановке сердца и смерти. При резком погружении в ледяную воду происходит рефлекторное сужение сосудов, что и может стать причиной сердечно-сосудистой катастрофы. К тому же противопоказанием являются многие хронические заболевания, характерные для пожилого возраста, например, болезни головного мозга и сердечно-сосудистой системы, эндокринные заболевания, артериальная гипертензия и т.д. Кроме того, пребывание организма в подобных экстремальных условиях может дать чрезмерную нагрузку на иммунную систему и стать причиной развития ОРВИ, гриппа, пневмонии и других инфекционных заболеваний. Купание в проруби после 60-ти лет допустимо только, если человек полностью здоров и не первый год практикует так называемое моржевание. И даже в этом случае нырять в ледяную воду стоит лишь в специально оборудованных для этого местах в присутствии медиков и сотрудников МЧС
Н.И. Пирогова, врач-физиотерапевт, врач ЛФК Александр Мелия предупредил, что в пожилом возрасте ныряние в прорубь без соответствующей подготовки может привести к остановке сердца и смерти. При резком погружении в ледяную воду происходит рефлекторное сужение сосудов, что и может стать причиной сердечно-сосудистой катастрофы. К тому же противопоказанием являются многие хронические заболевания, характерные для пожилого возраста, например, болезни головного мозга и сердечно-сосудистой системы, эндокринные заболевания, артериальная гипертензия и т.д. Кроме того, пребывание организма в подобных экстремальных условиях может дать чрезмерную нагрузку на иммунную систему и стать причиной развития ОРВИ, гриппа, пневмонии и других инфекционных заболеваний. Купание в проруби после 60-ти лет допустимо только, если человек полностью здоров и не первый год практикует так называемое моржевание. И даже в этом случае нырять в ледяную воду стоит лишь в специально оборудованных для этого местах в присутствии медиков и сотрудников МЧС
Заместитель директора по гериатрии Российского геронтологического научно-клинического центра РНИМУ им. Н.И. Пирогова, доктор медицинских наук, главный гериатр Департамента здравоохранения Москвы Надежда Рунихина отметила высокий риск переохлаждения у пожилых людей при купании в проруби.
Н.И. Пирогова, доктор медицинских наук, главный гериатр Департамента здравоохранения Москвы Надежда Рунихина отметила высокий риск переохлаждения у пожилых людей при купании в проруби.
В пожилом и особенно старческом возрасте ухудшаются различные механизмы терморегуляции, контроль за температурой тела может нарушиться в результате развития некоторых возраст-ассоциированных заболеваний, например, атеросклероза периферических сосудов, эндокринных нарушений, саркопении, недостаточности питания, снижения функциональной и физической активности, ухудшения когнитивного статуса.
Пожилые люди подвержены повышенному риску нарушений терморегуляции в результате снижения сердечной функции и уменьшения внутрисосудистого объема. Нарушение мышечной функции, снижение метаболизма или длительное воздействие низких температур окружающей среды могут привести к переохлаждению, потребление кислорода и активность ферментов снижается, ухудшаются когнитивные функции и может развиться делирий.
Отгрузки российского СПГ из Сабетты повысились в 2021 году
Отгрузки сжиженного природного газа с предприятия «Ямал СПГ», расположенного в арктическом порту Сабетта, в 2021 г. увеличились, чему способствовали пусконаладочные работы на четвертой линии завода.
По данным аналитического сервиса Vortexa, в 2021 г. отгрузки СПГ из Сабетты составили 265 партий, что соответствует примерно 19,3 млн т. По информации Argus, из этого объема девять партий или около 660 тыс. т были отправлены в порт Мурманск для перегрузки на другие суда. Тем временем Уральское таможенное управление сообщило 19 января, что в 2021 г. оформило документы на экспорт из Сабетты 18,7 млн т или 257 танкерных партий СПГ.
По оценке Argus, на рынок Азиатского-Тихоокеанского региона (АТР) в прошлом году поступило 36% всех отгруженных с завода «Ямал СПГ» партий, тогда как в 2020 г. доля таких отгрузок составила 32%. По данным участников рынка, более половины из поступивших в АТР партий ямальского СПГ в прошлом году было перевезено прямыми рейсами – преимущественно по Северному морскому пути, хотя также было совершено несколько рейсов через Суэцкий канал или по маршруту вокруг Мыса Доброй Надежды. Остальные предназначенные для стран АТР партии СПГ были доставлены покупателям после промежуточной перевалки в Мурманске либо в портах Бельгии.
Остальные предназначенные для стран АТР партии СПГ были доставлены покупателям после промежуточной перевалки в Мурманске либо в портах Бельгии.
«Новатэк», являющийся основным акционером завода «Ямал СПГ», пока не раскрыл информацию о производстве СПГ на предприятии и объемах отгрузок продукции. Согласно опубликованным ранее данным «Новатэка», в 2020 г. отгрузки СПГ из Сабетты составили 18,6 млн т или 255 танкерных партий, а в 2019 г. — 18,4 млн т или 253 партии.
Четвертая линия на заводе «Ямал СПГ» начала работу еще весной 2021 г., но «Новатэк» несколько раз останавливал ее для отладки работы оборудования. К началу IV квартала линия так и не достигла проектной мощности. В ноябре руководство «Новатэка» сообщало, что четвертая линия в целом по итогам 2021 г. произведет около 200 тыс. т СПГ, а на полную загрузку выйдет только в 2022 г.
Мощность четвертой линии по производству СПГ составляет около 940 тыс. т/год. Сжижение газа на этой линии осуществляется по технологии «Арктический каскад», которая запатентована «Новатэком» и применяется впервые.
Остальные три линии завода «Ямал СПГ» имеют мощность 5,5 млн т/год каждая, сжижение газа на них производится по технологии американской компании Air Products. СПГ со всех четырех линий завода поступает в общее хранилище.
С вводом в строй четвертой линии «Ямал СПГ» номинальная мощность завода по выпуску СПГ превысила 17,4 млн т/год по сравнению с прежними 16,5 млн т/год. Но в предыдущие несколько лет завод «Ямал СПГ» уже производил более 18 млн т/год СПГ, поскольку работал с загрузкой выше 100%. В июне 2021 г. председатель правления «Новатэка» Леонид Михельсон заявлял, что после окончательного ввода в строй четвертой очереди завод «Ямал СПГ» сможет производить более 20 млн т/год сжиженного газа.
«Новатэк» владеет 50,1% акций «Ямал СПГ». Остальными акционерами являются Total (20%), китайская CNPC (20%) и китайский инвестиционный Фонд шелкового пути (9,9%). Помимо сжиженного природного газа, завод «Ямал СПГ» выпускает газовый конденсат. Уральское таможенное управление в 2021 г. оформило документы на экспорт 20 танкерных партий конденсата общим объемом 827 тыс. т.
оформило документы на экспорт 20 танкерных партий конденсата общим объемом 827 тыс. т.
Передача сигналов Wnt/Fz и цитоскелет: потенциальная роль в онкогенезе
Davidson G, Wu W, Shen J, et al . Казеинкиназа 1γ связывает активацию рецептора Wnt с передачей цитоплазматического сигнала. Природа 2005; 438 :867–872.
КАС Статья пабмед Google Scholar
Цзэн Х, Тамай К., Добл Б., и др. . Двойной киназный механизм фосфорилирования и активации корецептора Wnt. Природа 2005; 438 :873–877.
КАС Статья пабмед ПабМед Центральный Google Scholar
Moon RT, Kohn AD, Ferrari GVD, Kaykas A . WNT и передача сигналов β-катенина: заболевания и методы лечения. Nat Rev Genet 2004; 5 :691–701.
КАС Статья пабмед Google Scholar
Толвински Н. С., Вишаус Э. .Переосмысление сигнализации WNT. Тенденции Genet 2004; 20 :177–181.
С., Вишаус Э. .Переосмысление сигнализации WNT. Тенденции Genet 2004; 20 :177–181.
КАС Статья пабмед Google Scholar
Нельсон В.Дж., Нассе Р. Конвергенция путей Wnt, β-катенина и кадгерина. Наука 2004; 303 :1483–1487.
КАС Статья пабмед ПабМед Центральный Google Scholar
Хе Х, Семенов М, Тамай К, Цзэн Х .Белки 5 и 6, связанные с рецептором ЛПНП, в передаче сигналов Wnt/β-катенина: стрелки указывают путь. Разработка 2004; 131 :1663–1677.
КАС Статья пабмед Google Scholar
Торбан Э., Кор С., Грос П. . Van Gogh-like2 (косоглазие) и его роль в плоскостной клеточной полярности и конвергентном расширении у позвоночных. Тенденции Genet 2004; 20 :570–577.
КАС Статья пабмед Google Scholar
Veeman MT, Axelrod JD, Moon RT . Второй канон: функции и механизмы независимой от β-catenin передачи сигналов Wnt. Dev Cell 2003; 5 :367–377.
Второй канон: функции и механизмы независимой от β-catenin передачи сигналов Wnt. Dev Cell 2003; 5 :367–377.
КАС Статья пабмед Google Scholar
Уоллингфорд Дж.Б., Хабас Р. Биология развития Dishevelled: загадочный белок, управляющий клеточной судьбой и клеточной полярностью. Разработка 2005; 132 :4421–4436.
КАС Статья пабмед Google Scholar
Ким Г.Х., Хан Дж.К.JNK и ROKα функционируют в неканоническом сигнальном пути Wnt/RhoA, чтобы регулировать движения конвергентного растяжения Xenopus. Дев Дин 2005; 232 :958–968.
КАС Статья пабмед Google Scholar
Даркен Р.С., Скола А.М., Ракеман А.С., и др. . Ген планарной полярности косоглазие регулирует движения конвергентного растяжения у Xenopus. EMBO J 2002; 21 :976–985.
КАС Статья пабмед ПабМед Центральный Google Scholar
Гото Т., Келлер Р. Ген планарной клеточной полярности косоглазие регулирует конвергенцию и растяжение, а также закрытие нервных складок у Xenopus. Дев Биол 2002; 247 :165–181.
КАС Статья пабмед Google Scholar
Парк М, Мун РТ . Ген планарной клеточной полярности stbm регулирует клеточное поведение и клеточную судьбу у эмбрионов позвоночных. Nat Cell Biol 2002; 4 :20–25.
КАС Статья пабмед Google Scholar
Кибар З., Воган К.Дж., Гроулкс Н., и др. . Ltap, гомолог Drosophila Strabismus/Van Gogh у млекопитающих, изменен в мутанте Loop-tail нервной трубки мыши. Нат Жене 2001; 28 :251–255.
КАС Статья пабмед Google Scholar
Каррейра-Барбоза Ф. , Конча М.Л., Такеучи М., и др. .Prickle 1 регулирует движения клеток во время гаструляции и миграции нейронов у рыбок данио. Разработка 2003; 130 :4037–4046.
, Конча М.Л., Такеучи М., и др. .Prickle 1 регулирует движения клеток во время гаструляции и миграции нейронов у рыбок данио. Разработка 2003; 130 :4037–4046.
КАС Статья пабмед Google Scholar
Такеучи М., Накабаяши Дж., Сакагути Т., и др. . Родственный с шипами ген у позвоночных необходим для движений гаструляционных клеток. Карр Биол 2003; 13 :674–679.
КАС Статья пабмед Google Scholar
Veeman MT, Slusarski DC, Kaykas A, Louie SH, Moon RT .Рыбка данио Prickle, модулятор неканонической передачи сигналов Wnt/Fz, регулирует гаструляционные движения. Карр Биол 2003; 13 :680–685.
КАС Статья пабмед Google Scholar
Куртин Дж.А., Квинт Э., Ципури В., и др. . Мутация Celsr1 нарушает планарную полярность волосковых клеток внутреннего уха и вызывает серьезные дефекты нервной трубки у мышей. Карр Биол 2003; 13 :1129–1133.
Карр Биол 2003; 13 :1129–1133.
КАС Статья пабмед Google Scholar
Цянь Д., Джонс С., Рзадзинска А., и др. . Wnt5a участвует в регуляции плоскостной клеточной полярности у мышей. Дев Биол 2007; 306 :121–133.
КАС Статья пабмед ПабМед Центральный Google Scholar
Kohn AD, Moon RT . Передача сигналов Wnt и кальция: пути, независимые от β-катенина. Клеточный кальций 2005; 38 :439–446.
КАС Статья пабмед Google Scholar
Слюсарский округ Колумбия, Пелегри Ф . Передача сигналов кальция в формировании эмбрионального паттерна и морфогенезе позвоночных. Дев Биол 2007; 307 :1–13.
КАС Статья пабмед ПабМед Центральный Google Scholar
Слюсарский округ Колумбия, Ян-Снайдер Дж. , Буса В.Б., Мун РТ .Модуляция эмбриональной внутриклеточной передачи сигналов Ca 2+ с помощью Wnt-5A. Дев Биол 1997; 182 :114–120.
, Буса В.Б., Мун РТ .Модуляция эмбриональной внутриклеточной передачи сигналов Ca 2+ с помощью Wnt-5A. Дев Биол 1997; 182 :114–120.
КАС Статья пабмед Google Scholar
Sheldahl LC, Park M, Malbon CC, Moon RT . Протеинкиназа С по-разному стимулируется гомологами Wnt и Frizzled зависимым от G-белка образом. Карр Биол 1999; 9 :695–698.
КАС Статья пабмед Google Scholar
Топчевски Дж., Сепич Д.С., Майерс Д.С., и др. .Glypican knypek рыбок данио контролирует полярность клеток во время гаструляционных движений конвергентного растяжения. Dev Cell 2001; 1 :251–264.
КАС Статья пабмед Google Scholar
Хикаса Х., Шибата М., Хиратани И., Тайра М. . Тирозинкиназа рецептора Xenopus Xror2 модулирует морфогенетические движения аксиальной мезодермы и нейроэктодермы посредством передачи сигналов Wnt. Разработка 2002; 129 :5227–5239.
Разработка 2002; 129 :5227–5239.
КАС пабмед Google Scholar
Kuhl M, Sheldahl LC, Malbon CC, Moon RT . Ca 2+ /calmodulin-зависимая протеинкиназа II стимулируется гомологами Wnt и Frizzled и способствует судьбе вентральных клеток у Xenopus. J Biol Chem 2000; 275 :12701–12711.
КАС Статья пабмед Google Scholar
Гейзенберг С.П., Тада М., Раух Г.Дж., и др. .Silberblick/Wnt11 опосредует конвергентные движения растяжения во время гаструляции рыбок данио. Природа 2000; 405 :76–81.
КАС Статья пабмед Google Scholar
Gilland E, Miller AL, Karplus E, Baker R, Webb SE . Визуализация многоклеточных крупномасштабных ритмичных кальциевых волн во время гаструляции рыбок данио. Proc Natl Acad Sci USA 1999; 96 : 157–161.
КАС Статья пабмед Google Scholar
Уоллингфорд Дж. Б., Эвальд А. Дж., Харланд Р. М., Фрейзер С. Э. .Передача сигналов кальция во время конвергентного расширения у Xenopus. Карр Биол 2001; 11 :652–661.
КАС Статья пабмед Google Scholar
Westfall TA, Brimeyer R, Twedt J, и др. . Wnt-5/pipettail функционирует при формировании оси позвоночных как негативный регулятор активности Wnt/β-catenin. J Cell Biol 2003; 162 :889–898.
КАС Статья пабмед ПабМед Центральный Google Scholar
Килиан Б., Мансукоски Х., Барбоза Ф.К., и др. .Роль Ppt/Wnt5 в регуляции формы и движения клеток во время гаструляции рыбок данио. Мех Дев 2003; 120 :467–476.
КАС Статья пабмед Google Scholar
Weeraratna AT, Jiang Y, Hostetter G, и др. . Передача сигналов Wnt5a напрямую влияет на подвижность клеток и инвазию метастатической меланомы. Раковая ячейка 2002; 1 :279–288.
. Передача сигналов Wnt5a напрямую влияет на подвижность клеток и инвазию метастатической меланомы. Раковая ячейка 2002; 1 :279–288.
КАС Статья Google Scholar
Деймек Дж., Сафхольм А., Камп Н.С., Андерссон Т., Леандерссон К. .Активность NFAT, индуцированная Wnt-5a/Ca 2+ , противодействует передаче сигналов Wnt-5a/Yes-Cdc42-казеинкиназы 1α в эпителиальных клетках молочной железы человека. Mol Cell Biol 2006; 26 :6024–6036.
КАС Статья пабмед ПабМед Центральный Google Scholar
Ма Л, Ван ХИ. Подавление циклической GMP-зависимой протеинкиназы необходимо для пути Wnt/cGMP/Ca 2+ . J Biol Chem 2006; 281 :30990–31001.
КАС Статья пабмед Google Scholar
Крылова О, Вестник М.Дж., Салинас П. С. Dishavelled-1 регулирует стабильность микротрубочек: новая функция, опосредованная киназой-3β гликогенсинтазы. J Cell Biol 2000; 151 :83–94.
С. Dishavelled-1 регулирует стабильность микротрубочек: новая функция, опосредованная киназой-3β гликогенсинтазы. J Cell Biol 2000; 151 :83–94.
КАС Статья пабмед ПабМед Центральный Google Scholar
Чиани Л., Крылова О., Смолли М.Дж., Дейл Т.С., Салинас П.С.Расходящийся канонический путь передачи сигналов WNT регулирует динамику микротрубочек: растрепанные сигналы локально стабилизируют микротрубочки. J Cell Biol 2004; 164 :243–253.
КАС Статья пабмед ПабМед Центральный Google Scholar
Чиани Л., Салинас П. . N-концевая киназа c-Jun (JNK) взаимодействует с Gsk3β, чтобы регулировать стабильность микротрубочек, опосредованную Dishevelled. BMC Cell Biol 2007; 8 :27.
Артикул КАС пабмед ПабМед Центральный Google Scholar
Чанг Л. , Джонс Ю., Эллисман М.Х., Гольдштейн Л.С., Карин М. . JNK1 необходим для поддержания нейрональных микротрубочек и контролирует фосфорилирование белков, связанных с микротрубочками. Dev Cell 2003; 4 :521–533.
, Джонс Ю., Эллисман М.Х., Гольдштейн Л.С., Карин М. . JNK1 необходим для поддержания нейрональных микротрубочек и контролирует фосфорилирование белков, связанных с микротрубочками. Dev Cell 2003; 4 :521–533.
КАС Статья пабмед Google Scholar
Шлессингер К., Макманус Э.Дж., Холл А.Cdc42 и неканонические пути передачи сигнала Wnt взаимодействуют, чтобы способствовать клеточной полярности. J Cell Biol 2007; 178 :355–361.
КАС Статья пабмед ПабМед Центральный Google Scholar
Zhang X, Zhu J, Yang GY, и др. . Disheveled способствует дифференцировке аксонов, регулируя атипичную протеинкиназу C. Nat Cell Biol 2007; 9 :743–754.
КАС Статья пабмед Google Scholar
Россо С.Б., Суссман Д., Уиншоу-Борис А. , Салинас П.С.Передача сигналов Wnt через Dishavelled, Rac и JNK регулирует развитие дендритов. Nat Neurosci 2005; 8 :34–42.
, Салинас П.С.Передача сигналов Wnt через Dishavelled, Rac и JNK регулирует развитие дендритов. Nat Neurosci 2005; 8 :34–42.
КАС Статья пабмед Google Scholar
Фан С., Рамирес С.Х., Гарсия Т.М., Дьюхерст С. . Disheveled способствует росту нейритов в нейронах, дифференцирующихся в клетках нейробластомы 2A, через путь, зависимый от домена DIX. Brain Res Mol Brain Res 2004; 132 :38–50.
КАС Статья пабмед Google Scholar
Капеллуто ДГС, Кутателадзе Т.Г., Хабас Р., и др. . Домен DIX нацелен на растрепанные актиновые стрессовые волокна и везикулярные мембраны. Природа 2002; 419 :726–729.
КАС Статья пабмед ПабМед Центральный Google Scholar
Kinzler KW, Vogelstein B .Уроки наследственного колоректального рака. Сотовый 1996; 87 :159–170.
Сотовый 1996; 87 :159–170.
КАС Статья пабмед Google Scholar
Бьенц М, Клеверс Х . Связывание колоректального рака с передачей сигналов Wnt. Сотовый 2000; 103 :311–320.
КАС Статья пабмед Google Scholar
Кэдиган К.М., Нуссе Р. Передача сигналов Wnt: общая тема в развитии животных. Гены Дев 1997; 11 :3286–3305.
КАС Статья пабмед Google Scholar
Фодде Р., Смитс Р., Клеверс Х. . APC, сигнальная трансдукция и генетическая нестабильность при колоректальном раке. Nat Rev Рак 2001; 1 :55–67.
КАС Статья пабмед Google Scholar
Полакис П . Передача сигналов Wnt и рак. Гены Дев 2000; 14 :1837–1851.
КАС пабмед Google Scholar
Samowitz WS, Powers MD, Spirio LN, и др. . Мутации β-катенина чаще встречаются в небольших колоректальных аденомах, чем в более крупных аденомах и инвазивных карциномах. Рак Res 1999; 59 :1442–1444.
КАС пабмед Google Scholar
Мимори-Кийосуэ Ю., Сиина Н., Цукита С. .Белок аденоматозного полипоза толстой кишки (АПК) перемещается по микротрубочкам и концентрируется на их растущих концах в эпителиальных клетках. J Cell Biol 2000; 148 :505–518.
КАС Статья пабмед ПабМед Центральный Google Scholar
Мунэмицу С., Соуза Б., Мюллер О., и др. . Продукт гена APC связывается с микротрубочками in vivo и способствует их сборке in vitro . Рак Res 1994; 54 :3676–3681.
КАС пабмед Google Scholar
Su LK, Burrell M, Hill DE, и др. . APC связывается с новым белком EB1. Рак Res 1995; 55 :2972–2977.
КАС пабмед Google Scholar
Асхэм Дж. М., Монкур П., Маркхэм А. Ф., Моррисон Э. Э. . Регуляция и функция взаимодействия между белком-онкосупрессором APC и EB1. Онкоген 2000; 19 : 1950–1958.
КАС Статья пабмед Google Scholar
Джимбо Т., Кавасаки Ю., Кояма Р., и др. . Выявление связи между APC-супрессором опухоли и надсемейством кинезинов. Nat Cell Biol 2002; 4 :323–327.
КАС Статья пабмед Google Scholar
Мимори-Кийосуэ Ю., Шиина Н., Цукита С. .Динамическое поведение APC-связывающего белка EB1 на дистальных концах микротрубочек. Карр Биол 2000; 10 :865–868.
Карр Биол 2000; 10 :865–868.
КАС Статья пабмед Google Scholar
Wen Y, Eng CH, Schmoranzer J, и др. . EB1 и APC связываются с mDia, чтобы стабилизировать микротрубочки ниже Rho и способствовать миграции клеток. Nat Cell Biol 2004; 6 :820–830.
КАС Статья пабмед Google Scholar
Зумбрунн Дж., Киношита К., Хайман А.А., Натке И.С.Связывание белка аденоматозного полипоза кишечной палочки с микротрубочками увеличивает стабильность микротрубочек и регулируется фосфорилированием GSK3β. Карр Биол 2001; 11 :44–49.
КАС Статья пабмед Google Scholar
Etienne-Manneville S, Hall A . Cdc42 регулирует GSK-3β и аденоматозный полипоз толстой кишки, чтобы контролировать клеточную полярность. Природа 2003; 421 :753–756.
КАС Статья Google Scholar
Грохманн А., Таннебергер К., Альзнер А., Шнайкерт Дж., Беренс Дж. .AMER1 регулирует распределение APC-супрессора опухоли между микротрубочками и плазматической мембраной. J Cell Sci 2007; 120 :3738–3747.
КАС Статья пабмед Google Scholar
Натке И.С., Адамс К.Л., Полакис П., Селлин Дж.Х., Нельсон В.Дж. Белок-супрессор опухоли аденоматозного полипоза толстой кишки локализуется в участках плазматической мембраны, участвующих в активной миграции клеток. J Cell Biol 1996; 134 :165–179.
КАС Статья пабмед Google Scholar
Салинас ПК . Модуляция цитоскелета микротрубочек: роль расходящегося канонического пути Wnt. Trends Cell Biol 2007; 17 :333–342.
КАС Статья пабмед Google Scholar
Шлезингер А. , Шелтон К.А., Малуф Дж.Н., Менегини М., Бауэрман Б. Компоненты пути Wnt ориентируют митотическое веретено у ранних эмбрионов Caenorhabditis elegans, не требуя транскрипции гена в отвечающей клетке. Гены Дев 1999; 13 :2028–2038.
, Шелтон К.А., Малуф Дж.Н., Менегини М., Бауэрман Б. Компоненты пути Wnt ориентируют митотическое веретено у ранних эмбрионов Caenorhabditis elegans, не требуя транскрипции гена в отвечающей клетке. Гены Дев 1999; 13 :2028–2038.
КАС Статья пабмед ПабМед Центральный Google Scholar
Чжан Х., Скоп А.Р., Уайт Д.Г. Передача сигналов Src и Wnt регулирует накопление dynactin на границе клеток P2-EMS у эмбрионов C. elegans. J Cell Sci 2008; 121 :155–161.
КАС Статья пабмед Google Scholar
Лу Б., Рогирс Ф., Ян Л.И., Ян Ю.Н.Слипчивые соединения ингибируют асимметричное деление эпителия дрозофилы. Природа 2001; 409 :522–525.
КАС Статья Google Scholar
Purro SA, Ciani L, Hoyos-Flight M, и др. . Wnt регулирует поведение аксонов посредством изменения направления роста микротрубочек: новая роль аденоматозного полипоза толстой кишки. J Neurosci 2008; 28 :8644–8654.
J Neurosci 2008; 28 :8644–8654.
КАС Статья пабмед ПабМед Центральный Google Scholar
Ватанабэ Т., Ван С., Норитаке Дж., и др. .Взаимодействие с IQGAP1 связывает APC с Rac1, Cdc42 и актиновыми филаментами во время поляризации и миграции клеток. Dev Cell 2004; 7 :871–883.
КАС Статья пабмед Google Scholar
Briggs MW, Sacks DB . IQGAP1 как интегратор сигналов: Ca 2+ , кальмодулин, Cdc42 и цитоскелет. FEBS Lett 2003; 542 :7–11.
КАС Статья пабмед Google Scholar
Башур А.М., Фуллертон А.Т., Харт М.Дж., Блум Г.С.IQGAP1, Rac- и Cdc42-связывающий белок, напрямую связывает и сшивает микрофиламенты. J Cell Biol 1997; 137 :1555–1566.
КАС Статья пабмед ПабМед Центральный Google Scholar
Фуката М. , Ватанабэ Т., Норитаке Дж., и др. . Rac1 и Cdc42 захватывают микротрубочки через IQGAP1 и CLIP-170. Сотовый 2002; 109 :873–885.
, Ватанабэ Т., Норитаке Дж., и др. . Rac1 и Cdc42 захватывают микротрубочки через IQGAP1 и CLIP-170. Сотовый 2002; 109 :873–885.
КАС Статья пабмед Google Scholar
Кавасаки Ю., Сенда Т., Ишидате Т., и др. .Asef, связь между супрессором опухоли APC и передачей сигналов G-белком. Наука 2000; 289 :1194–1197.
КАС Статья пабмед Google Scholar
Акияма Т., Кавасаки Ю. . Передача сигналов Wnt и актиновый цитоскелет. Онкоген 2006; 25 :7538–7544.
КАС Статья пабмед Google Scholar
Кавасаки Ю., Сато Р., Акияма Т. .Мутированные APC и Asef участвуют в миграции клеток колоректальной опухоли. Nat Cell Biol 2003; 5 :211–215.
КАС Статья пабмед Google Scholar
Надаулд Л. Д., Чидестер С., Шелтон Д.Н., и др. . Двойная роль аденоматозного полипоза толстой кишки в регуляции биосинтеза ретиноевой кислоты и Wnt во время развития глаза. Proc Natl Acad Sci USA 2006; 103 :13409–13414.
Д., Чидестер С., Шелтон Д.Н., и др. . Двойная роль аденоматозного полипоза толстой кишки в регуляции биосинтеза ретиноевой кислоты и Wnt во время развития глаза. Proc Natl Acad Sci USA 2006; 103 :13409–13414.
КАС Статья пабмед Google Scholar
Айзингер А.Л., Надаулд Л.Д., Шелтон Д.Н., и др. . Ген-супрессор опухоли аденоматозного полипоза толстой кишки регулирует экспрессию циклооксигеназы-2 по механизму, который включает ретиноевую кислоту. J Biol Chem 2006; 281 :20474–20482.
КАС Статья пабмед Google Scholar
Танэгасима К., Чжао Х., Давид И.Б.WGEF активирует Rho в пути Wnt-PCP и контролирует конвергентное расширение при гаструляции Xenopus. EMBO J 2008; 27 :606–617.
КАС Статья пабмед ПабМед Центральный Google Scholar
Ковар ДР . Молекулярные детали формин-опосредованной сборки актина. Curr Opin Cell Biol 2006; 18 :11–17.
Молекулярные детали формин-опосредованной сборки актина. Curr Opin Cell Biol 2006; 18 :11–17.
КАС Статья пабмед Google Scholar
Хиггс Х.Н.Форминовые белки: доменный подход. Trends Biochem Sci 2005; 30 :342–353.
КАС Статья пабмед Google Scholar
Гуд Б.Л., Эк М.Дж. Механизм и функция форминов в контроле сборки актина. Annu Rev Biochem 2007: 76 : 593–627.
КАС Статья пабмед Google Scholar
Ватанабе Н., Хигасида К.Формины: процессивные капперы растущих актиновых филаментов. Exp Cell Res 2004; 301 :16–22.
КАС Статья пабмед Google Scholar
Зигмонд С.Х., Евангелиста М., Бун С., и др. . Форминовый дырявый кэп допускает удлинение в присутствии плотных кэпирующих белков. Карр Биол 2003; 13 :1820–1823.
Карр Биол 2003; 13 :1820–1823.
КАС Статья пабмед Google Scholar
Евангелиста М., Прюйн Д., Амберг Д.С., Бун С., Бретшер А. .Формины направляют Arp2/3-независимую сборку актиновых филаментов, чтобы поляризовать рост клеток у дрожжей. Nat Cell Biol 2002; 4 :32–41.
КАС Статья пабмед Google Scholar
Валлар Б.Дж., Альбертс А.С. Формины: активные каркасы, ремоделирующие цитоскелет. Trends Cell Biol 2003; 13 :435–446.
КАС Статья пабмед Google Scholar
Хиггс Х.Н., Петерсон К.Дж.Филогенетический анализ домена гомологии формина 2. Мол Биол Селл 2005; 16 :1–13.
КАС Статья пабмед ПабМед Центральный Google Scholar
Альбертс А. С. Идентификация ауторегуляторного домена ауторегуляторного домена белка гомологии формина, родственного карбоксиконцевому прозрачному белку. J Biol Chem 2001; 276 :2824–2830.
С. Идентификация ауторегуляторного домена ауторегуляторного домена белка гомологии формина, родственного карбоксиконцевому прозрачному белку. J Biol Chem 2001; 276 :2824–2830.
КАС Статья пабмед Google Scholar
Ламмерс М., Роуз Р., Скрима А., Виттингхофер А. .Регуляция mDia1 путем аутоингибирования и его высвобождение с помощью Rho*GTP. EMBO J 2005; 24 :4176–4187.
КАС Статья пабмед ПабМед Центральный Google Scholar
Хабас Р., Като Ю., Хе Х. Wnt/Frizzled активация Rho регулирует гаструляцию позвоночных и нуждается в новом белке Daam1, гомологичном Formin. Сотовый 2001; 107 :843–854.
КАС Статья пабмед Google Scholar
Лю В., Сато А., Хадка Д., и др. .Механизм активации форминового белка Daam1. Proc Natl Acad Sci USA 2008; 105 :210–215.
Proc Natl Acad Sci USA 2008; 105 :210–215.
КАС Статья пабмед Google Scholar
Марлоу Ф., Топчевски Дж., Сепич Д., Солница-Крезель Л. . Rho kinase 2 рыбок данио действует ниже Wnt11, опосредуя клеточную полярность и эффективное движение конвергенции и растяжения. Карр Биол 2002; 12 :876–884.
КАС Статья пабмед Google Scholar
Хабас R, He X .В: Уильям Э., Балч С. (ред.). Активация Rho и Rac посредством передачи сигналов Wnt/Frizzled. Нью-Йорк: Academic Press, 2006: 500–511.
Сато А., Хадка Д.К., Лю В., и др. . Профилин является эффектором для Daam1 в неканонической передаче сигналов Wnt и необходим для гаструляции позвоночных. Разработка 2006; 133 :4219–4231.
КАС Статья пабмед Google Scholar
Лу Дж. , Мэн В., Пой Ф., и др. .Структура домена Fh3 Daam1: значение для регуляции формином сборки актина. Дж Мол Биол 2007; 369 : 1258–1269.
, Мэн В., Пой Ф., и др. .Структура домена Fh3 Daam1: значение для регуляции формином сборки актина. Дж Мол Биол 2007; 369 : 1258–1269.
КАС Статья пабмед ПабМед Центральный Google Scholar
Аспенстром П., Рихнау Н., Йоханссон А.С. Связанный с diaphanous формин DAAM1 сотрудничает с Rho GTPases RhoA и Cdc42, CIP4 и Src в регуляции клеточного морфогенеза и динамики актина. Exp Cell Res 2006; 312 :2180–2194.
Артикул КАС пабмед Google Scholar
Кида Ю.С., Сато Т., Миясака К.Ю., Суто А., Огура Т. . Daam1 регулирует эндоцитоз EphB во время конвергентного удлинения хорды рыбок данио. Proc Natl Acad Sci USA 2007; 104 :6708–6713.
КАС Статья пабмед Google Scholar
Лоуренсон И.Д., Виммер-Клейкамп С. Х., Лок П., и др. .Эфрин-А5 индуцирует округление, вздутие и деадгезию клеток 293T, экспрессирующих EphA3, и клеток меланомы с помощью CrkII и Rho-опосредованной передачи сигналов. J Cell Sci 2002; 115 (часть 5): 1059–1072.
Х., Лок П., и др. .Эфрин-А5 индуцирует округление, вздутие и деадгезию клеток 293T, экспрессирующих EphA3, и клеток меланомы с помощью CrkII и Rho-опосредованной передачи сигналов. J Cell Sci 2002; 115 (часть 5): 1059–1072.
КАС пабмед Google Scholar
Батле Э., Хендерсон Дж.Т., Бехтель Х., и др. . β-catenin и TCF опосредуют позиционирование клеток в кишечном эпителии, контролируя экспрессию EphB/ephrinB. Сотовый 2002; 111 :251–263.
КАС Статья пабмед Google Scholar
Го Д.Л., Чжан Дж., Юэн С.Т., и др. . Снижение экспрессии EphB2, которое соответствует инвазии и метастазированию колоректальных опухолей. Канцерогенез 2006; 27 :454–464.
КАС Статья пабмед Google Scholar
Clevers H, Batlle E . Рецепторы EphB/EphrinB и передача сигналов Wnt при колоректальном раке. Рак Res 2006; 66 :2–5.
Рецепторы EphB/EphrinB и передача сигналов Wnt при колоректальном раке. Рак Res 2006; 66 :2–5.
КАС Статья пабмед Google Scholar
Zhu S, Liu L, Korzh V, Gong Z, Low BC . RhoA действует ниже Wnt5 и Wnt11, регулируя движения конвергенции и растяжения путем вовлечения эффекторов Rho Kinase и Diaphanous: использование рыбок данио in vivo в качестве модели для передачи сигналов GTPase. Сотовый сигнал 2006; 18 :359–372.
КАС Статья пабмед Google Scholar
Лай С.Л., Чанг С.Н., Ван П.Дж., Ли С.Дж.Rho опосредует цитокинез и эпиболию через ROCK у рыбок данио. Мол Репрод Дев 2005; 71 :186–196.
КАС Статья пабмед Google Scholar
Каррейра С., Гудолл Дж., Денат Л., и др. . Mitf-регуляция Dia1 контролирует пролиферацию и инвазивность меланомы. Гены Дев 2006; 20 :3426–3439.
Гены Дев 2006; 20 :3426–3439.
КАС Статья пабмед ПабМед Центральный Google Scholar
Preudhomme C, Roumier C, Hildebrand MP, и др. .Неслучайные реаранжировки 4p13 гена RhoH/TTF, кодирующего GTP-связывающий белок, при неходжкинской лимфоме и множественной миеломе. Онкоген 2000; 19 :2023–2032.
КАС Статья пабмед Google Scholar
Reuther GW, Lambert QT, Booden MA, и др. . Связанный с лейкемией фактор обмена гуаниновых нуклеотидов Rho, белок семейства Dbl, обнаруженный мутировавшим при лейкемии, вызывает трансформацию путем активации RhoA. J Biol Chem 2001; 276 :27145–27151.
КАС Статья пабмед Google Scholar
Курлас П.Дж., Страут М.П., Бекнелл Б., и др. . Идентификация гена 11q23, кодирующего фактор обмена гуаниновых нуклеотидов: свидетельство его слияния с MLL при остром миелоидном лейкозе. Proc Natl Acad Sci USA 2000; 97 :2145–2150.
Proc Natl Acad Sci USA 2000; 97 :2145–2150.
КАС Статья пабмед Google Scholar
Кларк Э.А., Голуб Т.Р., Ландер Э.С., Хайнс Р.О.Геномный анализ метастазов показывает существенную роль RhoC. Природа 2000; 406 :532–535.
КАС Статья пабмед Google Scholar
Аснар С., Фернандес-Валерон П., Эспина С., Лакаль Х.К. Rho GTPases: потенциальные кандидаты для противоопухолевой терапии. Рак Летт 2004; 206 :181–191.
КАС Статья пабмед Google Scholar
Гомес дель П.Т., Бенита С.А., Валерон П.Ф., Эспина С., Лакаль Дж.К.Экспрессия Rho GTPase в онкогенезе: свидетельство значимой связи. Bioessays 2005; 27 :602–613.
Артикул КАС Google Scholar
Жаулиак С. , Лопес-Родригес С., Шоу Л.М., и др. . Роль факторов транскрипции NFAT в инвазии интегрин-опосредованной карциномы. Nat Cell Biol 2002; 4 :540–544.
, Лопес-Родригес С., Шоу Л.М., и др. . Роль факторов транскрипции NFAT в инвазии интегрин-опосредованной карциномы. Nat Cell Biol 2002; 4 :540–544.
КАС Статья пабмед Google Scholar
Йоэли-Лернер М., Ю Г.К., Рабиновиц И., и др. .Akt блокирует подвижность и инвазию клеток рака молочной железы через фактор транскрипции NFAT. Мол Ячейка 2005; 20 :539–550.
КАС Статья пабмед Google Scholar
Коттрелл С., Бикнелл Д., Какламанис Л., Бодмер В.Ф. Молекулярный анализ мутаций APC при семейном аденоматозном полипозе и спорадических карциномах толстой кишки. Ланцет 1992; 340 :626–630.
КАС Статья пабмед Google Scholar
Пауэлл С.М., Зилз Н., Бизер-Барклай Ю., и др. .Мутации APC возникают на ранних стадиях колоректального онкогенеза. Природа 1992; 359 :235–237.
Природа 1992; 359 :235–237.
КАС Статья пабмед ПабМед Центральный Google Scholar
Морин П.Дж., Спаркс А.Б., Коринек В., и др. . Активация передачи сигналов β-катенин-Tcf при раке толстой кишки за счет мутаций в β-катенине или APC. Наука 1997; 275 :1787–1790.
КАС Статья Google Scholar
Лю С., Като Ю., Чжан З., и др. .β-Trcp связывает фосфорилирование-деградацию β-катенина и регулирует формирование оси Xenopus. Proc Natl Acad Sci USA 1999; 96 :6273–6278.
КАС Статья пабмед Google Scholar
Лю С., Ли Ю., Семенов М., и др. . Контроль фосфорилирования/деградации β-катенина с помощью двойного киназного механизма. Сотовый 2002; 108 :837–847.
КАС Статья пабмед Google Scholar
Амит С. , Хацубай А., Бирман Ю., и др. .Опосредованное аксином CKI фосфорилирование β-катенина по Ser 45: молекулярный переключатель пути Wnt. Гены Дев 2002; 16 :1066–1076.
, Хацубай А., Бирман Ю., и др. .Опосредованное аксином CKI фосфорилирование β-катенина по Ser 45: молекулярный переключатель пути Wnt. Гены Дев 2002; 16 :1066–1076.
КАС Статья пабмед ПабМед Центральный Google Scholar
Брембек Ф. Х., Росарио М., Бирчмайер В. . Уравновешивание клеточной адгезии и передачи сигналов Wnt, ключевая роль β-катенина. Curr Opin Genet Dev 2006; 16 :51–59.
КАС Статья пабмед Google Scholar
Роура С., Миравет С., Пьедра Дж., де Эррерос АГ, Дунах М. .Регуляция ассоциации E-кадгерин/катенин посредством фосфорилирования тирозина. J Biol Chem 1999; 274 :36734–36740.
КАС Статья пабмед Google Scholar
Пьедра Дж., Миравет С., Кастано Дж., и др. . Ассоциированные с катенином р120 тирозинкиназы Fer и Fyn регулируют фосфорилирование β-катенина Tyr-142 и взаимодействие β-катенин-α-катенин. Мол Селл Биол 2003; 23 :2287–2297.
Мол Селл Биол 2003; 23 :2287–2297.
КАС Статья пабмед ПабМед Центральный Google Scholar
Аберле Х., Шварц Х., Хошуецки Х., Кемлер Р. . Единичные аминокислотные замены в белках семейства генов броненосцев отменяют их связывание с α-катенином. J Biol Chem 1996; 271 :1520–1526.
КАС Статья пабмед Google Scholar
Покутта С., Вайс Висконсин.Структура области димеризации и связывания β-катенина α-катенина. Молекулярная ячейка 2000/3; 5 :533–543.
КАС Статья пабмед Google Scholar
Пьедра Дж., Мартинес Д., Кастано Дж., и др. . Регуляция структуры и активности β-катенина фосфорилированием тирозина. Журнал биологической химии 2001/6/1; 276 :20436–20443.
КАС Статья пабмед Google Scholar
Castano J, Raurell I, Piedra JA, и др. .N- и С-концевые хвосты β-катенина модулируют скоординированное связывание белков слипчивых соединений с β-катенином. J Biol Chem 2002; 277 :31541–31550.
.N- и С-концевые хвосты β-катенина модулируют скоординированное связывание белков слипчивых соединений с β-катенином. J Biol Chem 2002; 277 :31541–31550.
КАС Статья пабмед Google Scholar
Брембек Ф.Х., Шварц-Ромонд Т., Баккерс Дж., и др. . Существенная роль BCL9-2 в переключении между адгезивной и транскрипционной функциями β-катенина. Гены Дев 2004; 18 :2225–2230.
КАС Статья пабмед ПабМед Центральный Google Scholar
Graveel C, Su Y, Koeman J, и др. . Активирующие мутации Met создают уникальные профили опухолей у мышей с селективной дупликацией мутантного аллеля. Proc Natl Acad Sci USA 2004; 101 :17198–17203.
КАС Статья пабмед Google Scholar
Ван З., Шен Д., Парсонс Д. В., и др. .Мутационный анализ тирозинфосфатома при колоректальном раке. Наука 2004; 304 :1164–1166.
В., и др. .Мутационный анализ тирозинфосфатома при колоректальном раке. Наука 2004; 304 :1164–1166.
КАС Статья пабмед Google Scholar
Ли С . Передача сигналов киназы Src при лейкемии. Int J Biochem Cell Biol 2007; 39 :1483–1488.
КАС Статья пабмед ПабМед Центральный Google Scholar
Хильбиг А .Киназа Src и рак поджелудочной железы. Recent Results Cancer Res 2008; 177 :179–185.
КАС Статья пабмед Google Scholar
Саймонс М., Глой Дж., Ганнер А., и др. . Инверсин, генный продукт, мутировавший при нефронофтизе типа II, функционирует как молекулярный переключатель между сигнальными путями Wnt. Нат Жене 2005; 37 :537–543.
КАС Статья пабмед ПабМед Центральный Google Scholar
Орсулич С. , Хубер О., Аберле Х., Арнольд С., Кемлер Р. .Связывание E-кадгерина предотвращает ядерную локализацию β-катенина и трансактивацию, опосредованную β-катенином/LEF-1. J Cell Sci 1999; 112 (часть 8): 1237–1245.
, Хубер О., Аберле Х., Арнольд С., Кемлер Р. .Связывание E-кадгерина предотвращает ядерную локализацию β-катенина и трансактивацию, опосредованную β-катенином/LEF-1. J Cell Sci 1999; 112 (часть 8): 1237–1245.
КАС пабмед Google Scholar
Грэм Т.А., Уивер С., Мао Ф., Кимельман Д., Сюй В. . Кристаллическая структура комплекса β-катенин/Tcf. Сотовый 2000; 103 :885–896.
КАС Статья пабмед Google Scholar
фон Крис Дж.П., Уинбек Г., Асбранд С., и др. .Горячие точки в β-катенине для взаимодействия с LEF-1, кондуктином и APC. Nat Struct Biol 2000; 7 :800–807.
КАС Статья пабмед Google Scholar
Хубер А.Х., Вайс Висконсин . Структура комплекса β-катенин/Е-кадгерин и молекулярная основа распознавания различных лигандов β-катенином. Сотовый 2001; 105 :391–402.
Сотовый 2001; 105 :391–402.
КАС Статья пабмед Google Scholar
Стокингер А., Эгер А., Вольф Дж., Беуг Х., Фойснер Р. .Е-кадгерин регулирует рост клеток, модулируя зависимую от пролиферации транскрипционную активность β-катенина. J Cell Biol 2001; 154 :1185–1196.
КАС Статья пабмед ПабМед Центральный Google Scholar
Gottardi CJ, Wong E, Gumbiner BM . Е-кадгерин подавляет клеточную трансформацию путем ингибирования передачи сигналов β-катенина независимым от адгезии образом. J Cell Biol 2001; 153 :1049–1060.
КАС Статья пабмед ПабМед Центральный Google Scholar
Stigliano I, Puricelli L, Filmus J, и др. . Глипикан-3 регулирует миграцию, адгезию и организацию актинового цитоскелета в опухолевых клетках молочной железы посредством модуляции передачи сигналов Wnt. Рак молочной железы Res Treat 2009; 114 :251–262.
Рак молочной железы Res Treat 2009; 114 :251–262.
КАС Статья пабмед Google Scholar
Витце Э.С., Литман Э.С., Аргаст Г.М., Мун Р.Т., Ан Н.Г.Wnt5a контролирует клеточную полярность и направленное движение за счет поляризованного перераспределения рецепторов адгезии. Наука 2008; 320 :365–369.
КАС Статья пабмед ПабМед Центральный Google Scholar
Сатьямурти К., Мюрерс Дж., Мейер Ф., Патель Д., Херлин М. . Специфичные для Mel-CAM генетические супрессорные элементы ингибируют рост и инвазию меланомы за счет потери связи через щелевые соединения. Онкоген 2001; 20 :4676–4684.
КАС Статья пабмед Google Scholar
Чиджи М., Доусон С. Десмосомы: роль в развитии рака? Br J Рак 2007; 96 :1783–1787.
КАС Статья пабмед ПабМед Центральный Google Scholar
Грин К.Дж., Симпсон К.Л. Десмосомы: новый взгляд на классику. J Invest Dermatol 2007; 127 :2499–2515.
КАС Статья пабмед Google Scholar
Гециос С., Хуэн А.С., Грин К.Дж. Отработка прочности и гибкости десмосом. Nat Rev Mol Cell Biol 2004; 5 :271–281.
КАС Статья пабмед Google Scholar
Карновский А., Климковский М.В. Удвоение передней оси у Xenopus, вызванное сверхэкспрессией кадгерин-связывающего белка плакоглобина. Proc Natl Acad Sci USA 1995; 92 :4522–4526.
КАС Статья пабмед Google Scholar
Коначчи-Соррелл М.Е., Бен-Йедидиа Т., Штутман М., и др. . Nr-CAM является геном-мишенью пути β-катенина/LEF-1 при меланоме и раке толстой кишки, и его экспрессия усиливает подвижность и способствует онкогенезу. Гены Дев 2002; 16 :2058–2072.
Гены Дев 2002; 16 :2058–2072.
КАС Статья пабмед ПабМед Центральный Google Scholar
Мерриам Дж.М., Рубинштейн А.Б., Климковски М.В.Цитоплазматически закрепленный плакоглобин индуцирует WNT-подобный фенотип у Xenopus. Дев Биол 1997; 185 :67–81.
КАС Статья пабмед Google Scholar
Миравет С., Пьедра Дж., Миро Ф., и др. . Фактор транскрипции Tcf-4 содержит разные сайты связывания для β-катенина и плакоглобина. J Biol Chem 2002; 277 : 1884–1891.
КАС Статья пабмед Google Scholar
Миравет С., Пьедра Дж., Кастано Дж., и др. .Тирозиновое фосфорилирование плакоглобина вызывает противоположные эффекты на его ассоциацию с десмосомами и компонентами адгезионных соединений и модулирует β-катенин-опосредованную транскрипцию. Мол Селл Биол 2003; 23 :7391–7402.
Мол Селл Биол 2003; 23 :7391–7402.
КАС Статья пабмед ПабМед Центральный Google Scholar
Гарсия-Грас Э., Ломбарди Р., Джокондо М.Дж., и др. . Подавление канонической передачи сигналов Wnt/β-catenin ядерным плакоглобином резюмирует фенотип аритмогенной кардиомиопатии правого желудочка. J Clin Invest 2006; 116 :2012–2021.
КАС Статья пабмед ПабМед Центральный Google Scholar
Бидерманн К., Фогельсанг Х., Беккер И., и др. . Десмоглеин 2 экспрессируется аномально, а не мутирует при семейном и спорадическом раке желудка. J Патол 2005; 207 :199–206.
КАС Статья пабмед Google Scholar
Яширо М., Нисиока Н., Хиракава К. .Снижение экспрессии молекулы адгезии десмоглеина-2 связано с карциномой желудка диффузного типа. Евро J Рак 2006; 42 :2397–2403.
Евро J Рак 2006; 42 :2397–2403.
КАС Статья пабмед Google Scholar
Хан К., Харди Р., Хак А., и др. . Переключение десмоколлина при колоректальном раке. Br J Рак 2006; 95 :1367–1370.
КАС Статья пабмед ПабМед Центральный Google Scholar
Оширо М., Ким С., Возняк Р., и др. .Эпигенетическое молчание DSC3 является обычным явлением при раке молочной железы человека. Рак молочной железы Res 2005; 7 :R669–R680.
КАС Статья пабмед ПабМед Центральный Google Scholar
Кисимото Н., Цао Ю., Парк А., Сунь З. . Ген кистозной почки морского конька регулирует процессы, опосредованные ресничками, и пути Wnt. Dev Cell 2008; 14 :954–961.
КАС Статья пабмед Google Scholar
Росс А. Дж., Мэй-Симера Х., Эйхерс Э.Р., и др. .Нарушение цилиарных белков синдрома Барде-Бидля нарушает плоскую клеточную полярность у позвоночных. Нат Жене 2005; 37 :1135–1140.
Дж., Мэй-Симера Х., Эйхерс Э.Р., и др. .Нарушение цилиарных белков синдрома Барде-Бидля нарушает плоскую клеточную полярность у позвоночных. Нат Жене 2005; 37 :1135–1140.
КАС Статья пабмед Google Scholar
Blacque OE, Leroux MR . Синдром Барде-Бидля: формирующийся патомеханизм внутриклеточного транспорта. Cell Mol Life Sci 2006; 63 :2145–2161.
КАС Статья пабмед Google Scholar
Структура и функциональные свойства миметика норрина Wnt для передачи сигналов с помощью Frizzled4, Lrp5/6 и протеогликана
Основные вопросы, требующие решения :
1) Основной научной проблемой является вывод о том, что сайт R121W представляет собой сайт связывания LRP5/6.Этот вывод может быть правильным, просто он не полностью подтверждается данными. Действительно, в статье нет прямого измерения взаимодействия Norrin-Lrp5/6. Потеря передачи сигналов из-за мутации R121W также может быть связана с потерей взаимодействия ГАГ; этот сайт очень положительно заряжен. Основным объяснением того, что этот сайт является сайтом взаимодействия LRP5/6, по-видимому, является то, что он отличается от лучше охарактеризованного (авторами) сайта связывания ГАГ (там связывается SOS, а мутации нокаутируют связывание гепарина), но это нельзя исключать. что второй сайт также является сайтом связывания GAG.Если авторы хотят претендовать на идентификацию сайта связывания LRP, они должны, по крайней мере, показать, что R121W по-прежнему связывает колонку с гепарином так же прочно, как дикий тип, или, что еще лучше, показать, что R121W Norrin не связывает LRP-или смягчить или уточнить раздел (уже немного уточненный) об идентификации сайта связывания LRP .
Потеря передачи сигналов из-за мутации R121W также может быть связана с потерей взаимодействия ГАГ; этот сайт очень положительно заряжен. Основным объяснением того, что этот сайт является сайтом взаимодействия LRP5/6, по-видимому, является то, что он отличается от лучше охарактеризованного (авторами) сайта связывания ГАГ (там связывается SOS, а мутации нокаутируют связывание гепарина), но это нельзя исключать. что второй сайт также является сайтом связывания GAG.Если авторы хотят претендовать на идентификацию сайта связывания LRP, они должны, по крайней мере, показать, что R121W по-прежнему связывает колонку с гепарином так же прочно, как дикий тип, или, что еще лучше, показать, что R121W Norrin не связывает LRP-или смягчить или уточнить раздел (уже немного уточненный) об идентификации сайта связывания LRP .
Важно, чтобы авторы рассмотрели этот вопрос, тем более что он является одним из основных моментов расхождений с Ke et al. (как указывают авторы «Верификация сайта связывания ГАГ»). Авторы предполагают, что участок, предложенный Кеэл. др. быть сайтом связывания Lrp5/6 на норрине, на самом деле является сайтом связывания SOS/GAG, и их анализ выявил истинный сайт связывания Lrp5/6 (в частности, Ke et. al. измеряли связывание между Norrin и Lrp6).
Авторы предполагают, что участок, предложенный Кеэл. др. быть сайтом связывания Lrp5/6 на норрине, на самом деле является сайтом связывания SOS/GAG, и их анализ выявил истинный сайт связывания Lrp5/6 (в частности, Ke et. al. измеряли связывание между Norrin и Lrp6).
Мы благодарим рецензентов за то, что они призвали нас дополнительно подумать над этими важными вопросами. В ответ на их предложения мы провели значительный объем экспериментальной работы, результаты которой мы добавили в рукопись.Мы также пересмотрели текст, чтобы прояснить и подробно обсудить моменты совпадения и несоответствия между нашей работой и интерпретацией результатов, а также работой Ke et al. (Ке и др., 2013). Мы уделили особое внимание всем пунктам рукописи, чтобы убедиться, что наша интерпретация имеющихся в настоящее время данных является четко квалифицированной, когда это уместно.
Во-первых, мы провели эксперименты по связыванию SPR для взаимодействия норрина дикого типа с Lrp6 и добавили эти данные в рукопись (рис. 7E).В нашем анализе мы измерили сродство 2,87 мкМ к норрину, связывающемуся с Lrp6; Ке и др. сообщили о значении K d , равном 0,45 мкМ, на основе анализа гомологичной конкуренции AlphaScreen с использованием MBP-Norrin и Lrp6 (Ke et al., 2013). Мы отмечаем эту разницу в значениях аффинности (предположительно в результате очень разных форматов анализа) в обсуждении.
7E).В нашем анализе мы измерили сродство 2,87 мкМ к норрину, связывающемуся с Lrp6; Ке и др. сообщили о значении K d , равном 0,45 мкМ, на основе анализа гомологичной конкуренции AlphaScreen с использованием MBP-Norrin и Lrp6 (Ke et al., 2013). Мы отмечаем эту разницу в значениях аффинности (предположительно в результате очень разных форматов анализа) в обсуждении.
Во-вторых, как было предложено рецензентами, мы провели анализ связывания гепарина. Эти эксперименты показывают, что мутант норрина R121W по-прежнему связывается с гепарином с высокой аффинностью (рис. 8 — дополнение к рисунку 1C), подобно норрину дикого типа (рис. 7 — дополнение к рисунку 1A).Однако в результате проведения дополнительной продукции белка и функциональных анализов для мутанта Norrin R121W мы обнаружили, что этот белок имеет более низкую растворимость и стабильность, чем белок дикого типа. Мы добавили эту информацию в раздел «Картирование потенциального сайта связывания Lrp5/6 на норрине» и в легенду к рисунку 8 — дополнение к рисунку 1C. Такое поведение белка оказалось очень проблематичным, когда мы попытались провести измерения SPR мутанта Norrin R121W, связывающегося с Lrp6. Поэтому мы не смогли включить этот эксперимент в нашу редакцию.
Такое поведение белка оказалось очень проблематичным, когда мы попытались провести измерения SPR мутанта Norrin R121W, связывающегося с Lrp6. Поэтому мы не смогли включить этот эксперимент в нашу редакцию.
В-третьих, мы смягчили и оговорили утверждения в разделе «Картирование потенциального сайта связывания Lrp5/6 на норрине». Мы считаем, что пересмотренная рукопись предоставляет читателям сбалансированную оценку имеющихся данных о взаимодействии норрина и Lrp5/6, полученных из наших текущих исследований и ранее опубликованных Ke et al. В частности, Ke et al. предположили, что сайт связывания Lrp5/6 включает положительно заряженные остатки (Lys54, Arg107, Arg109, Arg115) и гидрофобные остатки (Leu52 и Tyr53).Они создали мутант MBP-Norrin K54E/R109E и обнаружили, что двойные мутации нарушают связывание Lrp6 в конкурентном анализе (MBP-Norrin против взаимодействия Lrp6 P1E1P2E2 и пептида DKK1). На самом деле эти результаты частично согласуются с предложенным нами сайтом связывания Lrp5/6, потому что наши текущие исследования также предполагают, что остаток Lys54 участвует в связывании Lrp5/6 (рис. 8). Однако наши структурные (рис. 5E) и функциональные анализы (рис. 7 и рис. 7 — дополнение 1 к рисунку) показывают, что Arg109, второй остаток мутанта MBP-Norrin K54E/R109E, способствует связыванию GAG, а не связыванию Lrp5/6.Теперь мы уточняем эти моменты в пересмотренной рукописи, помещая наши результаты в контекст предыдущих исследований с четкими обсуждениями в разделах «Проверка сайта связывания ГАГ» и «Картирование потенциального сайта связывания Lrp5/6 на норрине».
8). Однако наши структурные (рис. 5E) и функциональные анализы (рис. 7 и рис. 7 — дополнение 1 к рисунку) показывают, что Arg109, второй остаток мутанта MBP-Norrin K54E/R109E, способствует связыванию GAG, а не связыванию Lrp5/6.Теперь мы уточняем эти моменты в пересмотренной рукописи, помещая наши результаты в контекст предыдущих исследований с четкими обсуждениями в разделах «Проверка сайта связывания ГАГ» и «Картирование потенциального сайта связывания Lrp5/6 на норрине».
2) Ке и др. (2013) сообщают об экспрессии, кристаллизации и определении рентгеновской структуры слитого белка MBP-Norrin, который рефолдируется после экспрессии в E. coli . Таким образом, утверждения « технических трудностей при получении рекомбинантного норрина » в настоящей рукописи (Введение) преувеличены, если не вводят в заблуждение.Предыдущая работа была опубликована более года назад, и авторы уже давно представляют свою работу как конкурирующую с Ке, а не как построенную на Ке. Описания структуры Норрина в тексте и Рисунок 2 кажутся излишне избыточными с более ранней работой, и MS выиграет, сосредоточившись на сравнении их структур с более ранней структурой, чтобы выделить гибкие/консервативные области, например как показано на Рисунок 2-дополнение 3 D и E .
Описания структуры Норрина в тексте и Рисунок 2 кажутся излишне избыточными с более ранней работой, и MS выиграет, сосредоточившись на сравнении их структур с более ранней структурой, чтобы выделить гибкие/консервативные области, например как показано на Рисунок 2-дополнение 3 D и E .
Мы благодарим рецензентов за эти предложения. В ответ мы пересмотрели наш текст для производства белка Norrin (раздел «Введение»). Мы также сократили наше описание структуры норрина («Производство биологически активного норрина») и изменили рисунок 2 и рисунок 2 — приложение к рисунку 3, чтобы учесть комментарии рецензентов и полностью использовать доступность структуры из предыдущих исследований. Ке и др., (2013). Тем не менее, мы считаем, что наше нынешнее сокращенное описание структуры норрин-апо необходимо, чтобы сориентировать читателей перед разделами результатов сложных и обязательных исследований.
3) Утверждение « … количественных определений аффинности связывания норрина с различными CRD Fz » (Результаты) также вводит в заблуждение, поскольку оба Ke et al. (2013) и Xu et al. (2004) сообщают об однозначном наномолярном сродстве норрина к Fc-димеризованному Fz4 CRD . Аффинность взаимодействия между Norrin и Fz4 CRD значительно отличается в этом исследовании (приблизительно 1 микромоль по SPR) и в предыдущем исследовании Ke et.др. (~10 нМ с помощью Alphascreen и прибора Octet Red). Эти исследования могут немного отличаться тем, что они используют димеризованный CRD, что, по-видимому, является причиной того, что более ранние авторы измеряли более плотное связывание из-за авидности, но эти измерения, безусловно, количественные и даже, возможно, более физиологически релевантные, учитывая очевидную димеризацию Fz4 на клетках. Это следует отметить и обсудить возможные объяснения этого несоответствия .
(2013) и Xu et al. (2004) сообщают об однозначном наномолярном сродстве норрина к Fc-димеризованному Fz4 CRD . Аффинность взаимодействия между Norrin и Fz4 CRD значительно отличается в этом исследовании (приблизительно 1 микромоль по SPR) и в предыдущем исследовании Ke et.др. (~10 нМ с помощью Alphascreen и прибора Octet Red). Эти исследования могут немного отличаться тем, что они используют димеризованный CRD, что, по-видимому, является причиной того, что более ранние авторы измеряли более плотное связывание из-за авидности, но эти измерения, безусловно, количественные и даже, возможно, более физиологически релевантные, учитывая очевидную димеризацию Fz4 на клетках. Это следует отметить и обсудить возможные объяснения этого несоответствия .
Мы ценим комментарии рецензентов и учли их все в исправленной рукописи.Конкретно:
Ke et al. и Xu et al. действительно провели количественные измерения связывания норрина с Fc-димеризованным Fz4 CRD (Xu et al. , 2004, Ke et al., 2013), но они не обнаружили значительных взаимодействий, и, следовательно, измерить аффинность связывания норрина с CRD других рецепторов Frizzled. Из нашего структурного сравнения Fz4 CRD с Fz8 CRD (рис. 9D-F) мы предположили, что Fz8 CRD может иметь некоторую способность взаимодействовать с норрином.Поэтому мы продолжили исследования для измерения значений сродства связывания между норрином и CRD рецепторов Frizzled, включая Fz5 CRD , Fz7 CRD и Fz8 CRD . Мы согласны с комментарием рецензента о том, что наша первоначальная формулировка вводит в заблуждение, и изменили предложение «Поскольку не было проведено количественного определения сродства связывания норрина с различными CRD Fz» на «Чтобы определить сродство связывания норрина с различными CRD Fz». Фз».
, 2004, Ke et al., 2013), но они не обнаружили значительных взаимодействий, и, следовательно, измерить аффинность связывания норрина с CRD других рецепторов Frizzled. Из нашего структурного сравнения Fz4 CRD с Fz8 CRD (рис. 9D-F) мы предположили, что Fz8 CRD может иметь некоторую способность взаимодействовать с норрином.Поэтому мы продолжили исследования для измерения значений сродства связывания между норрином и CRD рецепторов Frizzled, включая Fz5 CRD , Fz7 CRD и Fz8 CRD . Мы согласны с комментарием рецензента о том, что наша первоначальная формулировка вводит в заблуждение, и изменили предложение «Поскольку не было проведено количественного определения сродства связывания норрина с различными CRD Fz» на «Чтобы определить сродство связывания норрина с различными CRD Fz». Фз».
Что касается различий в значениях сродства по сравнению с предыдущими исследованиями (Xu et al., 2004, Ke et al., 2013), мы первоначально кратко объяснили этот вопрос в легенде к рисунку 6 — приложение к рисунку 1. Однако рецензенты сделали несколько замечательных комментариев, которые мы использовали в нашем исправленном тексте; мы переместили наше предыдущее объяснение из подписи к рисунку 6 — приложение к рисунку 1 в раздел «Обсуждение» и теперь обсудим его более подробно.
4) Авторы показывают, что мутанты норрина в остатках, связывающих SOS, особенно мутант R107E/R109E/R115L, проявляют пониженную активность в анализе люциферазы для передачи сигналов Wnt ( Рисунок 6G ) и не способны связываться с гепарином ( Рисунок 6 — дополнение к рисунку 2Б).Могут ли авторы предоставить какие-либо доказательства того, что гликозаминогликаны (ГАГ) играют положительную роль в клеточных ответах на норрин, как предполагают и рисуют авторы во всех своих моделях? Физиологическую релевантность этого взаимодействия для передачи сигналов важно установить, т.к. это центральный аспект структуры .
Мы благодарим рецензентов за то, что они подняли этот вопрос. Мы действительно рассматривали возможность проведения дальнейших исследований для изучения роли HSPG в передаче сигналов Norrin. Мы согласны с рецензентами в том, что это интересный и важный вопрос для рассмотрения, однако мы считаем, что для того, чтобы сделать это в полном объеме, потребуется значительная работа, которая выходит за рамки нашего текущего исследования.Поэтому в ответ на этот комментарий:
Мы согласны с рецензентами в том, что это интересный и важный вопрос для рассмотрения, однако мы считаем, что для того, чтобы сделать это в полном объеме, потребуется значительная работа, которая выходит за рамки нашего текущего исследования.Поэтому в ответ на этот комментарий:
Во-первых, мы включили новые данные анализа связывания SPR (рис. 7F), чтобы показать, что мутант норрина (R107E/R109E/R115L) в сайте связывания ГАГ сохраняет способность связываться с Lrp6 P1E1P2E2 . Мы сделали новый рисунок 7, посвященный проверке сайта связывания ГАГ, и добавили дополнительное описание наших результатов в раздел «Проверка сайта связывания ГАГ».
Во-вторых, мы добавили больше описания и разъяснили наше обсуждение HSPG в разделе «Обсуждение».
В-третьих, мы приняли к сведению опасения рецензентов (в комментариях 4 и 5) и, поразмыслив, пришли к выводу, что наши схемы на различных рисунках могут дать читателю слишком упрощенное представление о возможных путях, которыми могут способствовать ГАГ. к сигнализации Норрин. Поэтому мы решили удалить старый рисунок 8G (карикатурная модель индуцированной норрином кластеризации рецепторов и образования сигналлосом) и рисунок 6, дополненный рисунком 2D.
к сигнализации Норрин. Поэтому мы решили удалить старый рисунок 8G (карикатурная модель индуцированной норрином кластеризации рецепторов и образования сигналлосом) и рисунок 6, дополненный рисунком 2D.
5) Авторы предполагают, что молекула SOS в структуре (и, предположительно, ГАГ в клетках; см. модель на рис. 6, дополнение 2) связывает взаимодействие между Norrin и Fz4 CRD .В качестве подтверждения они ссылаются на статью (Smallwood JBC 2007), в которой показано, что гепарин может повышать сродство между Norrin и Fz4 CRD примерно в 10 раз. Прямых экспериментальных испытаний этой модели не проводилось (за исключением трудно интерпретируемого эксперимента, согласно которому избыток SOS может снижать сигнальную эффективность норрина). Простым (но важным) тестом этой модели было бы выяснить, могут ли (1) SOS или гепарин усиливать сродство взаимодействия Norrin-Fz4 CRD в их анализе SPR и (2) могут ли мутации в предполагаемый сайт связывания SOS норрина (например,г. тройная мутация R107E/R109E/R115L, показанная на Рисунке 6, дополнение 2), может отменить этот эффект гепарина/SOS (хотя они мало влияют на исходное взаимодействие Norrin-Fz4 CRD ) .
тройная мутация R107E/R109E/R115L, показанная на Рисунке 6, дополнение 2), может отменить этот эффект гепарина/SOS (хотя они мало влияют на исходное взаимодействие Norrin-Fz4 CRD ) .
Мы ценим комментарии рецензентов относительно спекулятивного характера нашей модели роли молекулы SOS/GAG в качестве дополнительного компонента комплекса Norrin-Fz4 CRD . Мы провели серию дополнительных экспериментов, и хотя мы можем предоставить существенные дополнительные данные в поддержку особой природы сайта связывания ГАГ на норрине, мы не можем предоставить окончательные доказательства особой важности мостиковой роли в комплексообразовании.Соответственно, мы изменили баланс нашего обсуждения этого вопроса в исправленной рукописи. Конкретно:
Мы приносим свои извинения за неясность в нашем описании того, как избыток SOS может ингибировать передачу сигналов норрина, и поясняем наше обсуждение этого эксперимента в разделе «Обсуждение», ссылаясь на два примера, в которых было показано, что экзогенный гепарин и PG545 (миметик гепарансульфата) ингибируют передачу сигналов Wnt путем конкуренции с эндогенными HSPG за взаимодействия с лигандами и/или рецепторами (Ai et al. , 2003, Юнг и др., 2015). В попытке полностью ответить на комментарии рецензентов мы попытались измерить аффинность связывания норрина дикого типа или мутанта R107E/R109E/R115L с Fz4 CRD в присутствии SOS. Тем не менее, мы не смогли обнаружить последовательные, значительные изменения в аффинности с помощью нашего анализа SPR. По размышлении мы подозреваем, что, поскольку сайт связывания ГАГ охватывает норрин и Fz4 CRD (рис. 5E), ингибирующие эффекты отдельных связываний норрин-SOS и событий связывания Fz4 CRD –SOS блокируют образование норрин-SOS-Fz4 CRD. (аналогично нашему анализу ингибирования SOS; рисунок 7 — дополнение к рисунку 1D) может нейтрализовать любой стабилизирующий вклад в образование комплексов в этих измерениях in vitro.
, 2003, Юнг и др., 2015). В попытке полностью ответить на комментарии рецензентов мы попытались измерить аффинность связывания норрина дикого типа или мутанта R107E/R109E/R115L с Fz4 CRD в присутствии SOS. Тем не менее, мы не смогли обнаружить последовательные, значительные изменения в аффинности с помощью нашего анализа SPR. По размышлении мы подозреваем, что, поскольку сайт связывания ГАГ охватывает норрин и Fz4 CRD (рис. 5E), ингибирующие эффекты отдельных связываний норрин-SOS и событий связывания Fz4 CRD –SOS блокируют образование норрин-SOS-Fz4 CRD. (аналогично нашему анализу ингибирования SOS; рисунок 7 — дополнение к рисунку 1D) может нейтрализовать любой стабилизирующий вклад в образование комплексов в этих измерениях in vitro.
Следует отметить, что кристаллы нативного комплекса Norrin-Fz4 CRD росли только в присутствии SOS (рис. 4A) и что структурный анализ показывает молекулу SOS, охватывающую поверхность раздела между Norrin и Fz4 CRD (рис. 4E). . Это «связующее» действие предполагает механизм, с помощью которого можно объяснить наблюдение Смоллвуда и др. примерно 10-кратного усиления сродства между норрином и Fz4 CRD в присутствии гепарина.Поэтому мы считаем разумным сохранить некоторое упоминание о возможной значимости «моста SOS» для содействия формированию сигнальных комплексов в пересмотренной рукописи.
4E). . Это «связующее» действие предполагает механизм, с помощью которого можно объяснить наблюдение Смоллвуда и др. примерно 10-кратного усиления сродства между норрином и Fz4 CRD в присутствии гепарина.Поэтому мы считаем разумным сохранить некоторое упоминание о возможной значимости «моста SOS» для содействия формированию сигнальных комплексов в пересмотренной рукописи.
В нашей исходной рукописи мы сообщали, что мутации (R107E/R109E/R115L) в сайте связывания Norrin GAG не влияют на способность Norrin связывать Fz4 CRD (рис. 7D), но нарушают активацию передачи сигналов (рис. 7C). . Теперь мы провели дополнительные эксперименты, чтобы продемонстрировать образование комплекса Norrin R107E/R109E/R115L–Fz4 CRD (рис. 7 – дополнение к рисунку 1B) и взаимодействие с Lrp6 P1E1P2E2 (рис. 7F), а также подтвердили, что эти мутации нарушают работу гепарина. связывание (рис. 7B).В совокупности эти данные свидетельствуют о том, что сайт связывания GAG не зависит от сайтов связывания Fz4 и предполагаемых сайтов связывания Lrp5/6.
6) Авторы утверждают, что в силу того, что FzCRD являются мономерными при высокой концентрации в растворе « CRD Fz-рецепторов… не участвуют в димеризации рецепторов. ” Слабые взаимодействия в растворе все же могут иметь значение в двумерных мембранах, и даже если они не вносят значительного энергетического вклада, наблюдаемые контакты могут образовываться в димерах Fz (и влиять на характер взаимодействия), которые формируются в основном на основе благоприятных взаимодействий ТМ регионов .
Мы согласны с рецензентами. Мы не можем исключить возможность того, что CRD рецепторов Fz способствуют димеризации рецепторов в клеточной мембране, хотя наши исследования раствора in vitro показывают, что их взаимодействие очень слабое. Мы сейчас констатируем это при обсуждении этих результатов.
7) Любопытно, что единственный Fz4 CRD связывался с димером Norrin в отсутствие SOS. Есть ли у авторов какое-либо объяснение этой неожиданной стехиометрии (условия кристаллизации и т. д. ). )?
д. ). )?
Мы думаем, что это очень интересный вопрос. Конечно, такие результаты не уникальны, например, случай колониестимулирующего фактора-1 (CSF-1) и рецептора CSF-1, в котором димерный CSF-1 связан с одной молекулой CSF-1R в кристаллической структуре (Chen et al. др., 2008)
На самом деле мы исследовали, что может быть причиной нашей сложной структуры 2:1, но решили опустить наши выводы из оригинальной рукописи, потому что мы чувствовали, что они могут не представлять достаточно общего интереса.В нашем метилированном комплексе Norrin-Fz4 CRD (рисунок 4 — дополнение к рисунку 1B) мы обнаружили, что два остатка лизина, участвующие в интерфейсе Norrin-Fz4 CRD , диметилированы в незакомплексованной субъединице димера Norrin. Мы включили этот структурный анализ в рисунок 4 — приложение к рисунку 1C и добавили некоторые комментарии к разделу «Кристаллическая структура норрина в комплексе с Fz4 CRD ».
https://doi. org/10.7554/eLife.06554.028
org/10.7554/eLife.06554.028Регуляция передачи сигналов Wnt/β-катенина с помощью посттрансляционных модификаций | Cell & Bioscience
Kohn AD, Moon RT: передача сигналов Wnt и кальция: пути, независимые от β-катенина. Клеточный кальций. 2005, 38 (3–4): 439–446.
КАС пабмед Статья Google Scholar
De A: Сигнальный путь Wnt/Ca 2+ : краткий обзор. Акта Биохим Биофиз Син. 2011, 43 (10): 745-756. 10.1093/аббс/гмр079
КАС пабмед Статья Google Scholar
Veeman MT, Axelrod JD, Moon RT: Второй канон: функции и механизмы независимой от β-катенина передачи сигналов Wnt. Ячейка Дев. 2003, 5 (3): 367-377. 10.1016/С1534-5807(03)00266-1
КАС пабмед Статья Google Scholar
MacDonald BT, Tamai K, He X: Передача сигналов Wnt/β-катенина: компоненты, механизмы и заболевания. Ячейка Дев. 2009, 17 (1): 9-26. 10.1016/j.devcel.2009.06.016
Ячейка Дев. 2009, 17 (1): 9-26. 10.1016/j.devcel.2009.06.016
Центральный пабмед КАС пабмед Статья Google Scholar
Clevers H: Передача сигналов Wnt/β-катенина в развитии и заболевании. Клетка. 2006, 127 (3): 469-480. 10.1016/j.cell.2006.10.018
CAS пабмед Статья Google Scholar
Clevers H, Nusse R: Передача сигналов Wnt/β-катенина и заболевание. Клетка. 2012, 149 (6): 1192-1205. 10.1016/j.cell.2012.05.012
CAS пабмед Статья Google Scholar
Polakis P: Сигнализация Wnt и рак.Гены Дев. 2000, 14 (15): 1837-1851.
КАС пабмед Google Scholar
Кэдиган К.М., Пайфер М.: Передача сигналов Wnt от развития к заболеванию: понимание модельных систем. Колд Спринг Харб Перспект Биол. 2009, 1 (2): а002881.
Центральный пабмед пабмед Статья Google Scholar
Logan CY, Nusse R: Сигнальный путь Wnt в развитии и заболевании.Annu Rev Cell Dev Biol. 2004, 20 (1): 781-810. 10.1146/annurev.cellbio.20.010403.113126
CAS пабмед Статья Google Scholar
Luo J, Chen J, Deng ZL, Luo X, Song WX, Sharff KA, Tang N, Haydon RC, Luu HH, He TC: Передача сигналов Wnt и болезни человека: каковы терапевтические последствия?. Лаборатория Инвест. 2007, 87 (2): 97-103. 10.1038/labinvest.3700509
CAS пабмед Статья Google Scholar
Stamos JL, Weis WI: комплекс разрушения β-катенина. Колд Спринг Харб Перспект Биол. 2013, 5 (1): а007898.
Центральный пабмед пабмед Статья КАС Google Scholar
Liu C, Li Y, Semenov M, Han C, Baeg GH, Tan Y, Zhang Z, Lin X, He X: Контроль фосфорилирования/деградации β-катенина с помощью двойного киназного механизма. Клетка. 2002, 108 (6): 837-847. 10.1016/S0092-8674(02)00685-2
Клетка. 2002, 108 (6): 837-847. 10.1016/S0092-8674(02)00685-2
КАС пабмед Статья Google Scholar
Jiang J, Struhl G: Регуляция сигнальных путей Hedgehog и Wingless с помощью белка F-box/WD40-repeat Slimb. Природа. 1998, 391 (6666): 493-496. 10.1038/35154
КАС пабмед Статья Google Scholar
Liu C, Kato Y, Zhang Z, Do VM, Yankner BA, He X: β-Trcp связывает фосфорилирование-деградацию β-катенина и регулирует формирование оси Xenopus. Proc Natl Acad Sci U S A. 1999, 96 (11): 6273-6278. 10.1073/pnas.96.11.6273
PubMed Central КАС пабмед Статья Google Scholar
Cong F, Schweizer L, Varmus H: Wnt передает сигналы через плазматическую мембрану для активации пути β-катенина путем образования олигомеров, содержащих его рецепторы, Frizzled и LRP. Разработка. 2004, 131 (20): 5103-5115. 10.1242/dev.01318
2004, 131 (20): 5103-5115. 10.1242/dev.01318
CAS пабмед Статья Google Scholar
Hernández AR, Klein AM, Kirschner MW: Кинетические ответы β-катенина определяют сайты контроля Wnt. Наука. 2012, 338 (6112): 1337-1340. 10.1126/наука.1228734
PubMed Статья КАС Google Scholar
Kim SE, Huang H, Zhao M, Zhang X, Zhang A, Semonov MV, MacDonald BT, Zhang X, Abreu JG, Peng L, He X: Wnt стабилизация β-катенина раскрывает принципы рецептора морфогена -подмости в сборе. Наука.2013, 340 (6134): 867-870. 10.1126/science.1232389
PubMed Central КАС пабмед Статья Google Scholar
Li Vivian SW, Ng Ser S, Boersema Paul J, Low Teck Y, Karthaus Wouter R, Gerlach Jan P, Mohammed S, Heck Albert JR, Maurice Madelon M, Mahmoudi T, Clevers H: передача сигналов через Wnt ингибирование деградации β-катенина в интактном комплексе Axin1. Клетка. 2012, 149 (6): 1245-1256. 10.1016/j.cell.2012.05.002
Клетка. 2012, 149 (6): 1245-1256. 10.1016/j.cell.2012.05.002
КАС пабмед Статья Google Scholar
Wodarz A, Nusse R: Механизмы передачи сигналов Wnt в процессе развития. Annu Rev Cell Dev Biol. 1998, 14 (1): 59-88. 10.1146/annurev.cellbio.14.1.59
CAS пабмед Статья Google Scholar
Peifer M, Polakis P: Передача сигналов Wnt в онкогенезе и эмбриогенезе – взгляд вне ядра.Наука. 2000, 287 (5458): 1606-1609. 10.1126/наука.287.5458.1606
CAS пабмед Статья Google Scholar
Дерибе Ю.Л., Поусон Т., Дикич И.: Посттрансляционные модификации в интеграции сигналов. Nat Struct Mol Biol. 2010, 17 (6): 666-672. 10.1038/nsmb.1842
CAS пабмед Статья Google Scholar
Дженсен О.Н.: Интерпретация белкового языка с помощью протеомики. Nat Rev Mol Cell Biol. 2006, 7 (6): 391-403. 10.1038/nrm1939
Nat Rev Mol Cell Biol. 2006, 7 (6): 391-403. 10.1038/nrm1939
CAS пабмед Статья Google Scholar
Willert K, Nusse R: Белки Wnt. Колд Спринг Харб Перспект Биол. 2012, 4 (9): а007864.
Центральный пабмед пабмед Статья КАС Google Scholar
Ke J, Xu HE, Williams BO: Модификация липидов в структуре и функции Wnt.Карр Опин Липидол. 2013, 24 (2): 129-133. 10.1097/МОЛ.0б013е32835дф2бф
КАС пабмед Статья Google Scholar
Yanfeng WA, Tan C, Fagan RJ, Klein PS: Фосфорилирование frizzled-3. Дж. Биол. Хим. 2006, 281 (17): 11603-11609. 10.1074/джбк.М600713200
КАС пабмед Статья Google Scholar
Djiane A, Yogev S, Mlodzik M: Апикальные детерминанты aPKC и dPatj регулируют зависимую от Frizzled полярность планарных клеток в глазу дрозофилы. Клетка. 2005, 121 (4): 621-631. 10.1016/j.cell.2005.03.014
CAS пабмед Статья Google Scholar
Koo BK, Spit M, Jordens I, Low TY, Stange DE, van de Wetering M, van Es JH, Mohammed S, Heck AJR, Maurice MM, Clevers H: Супрессор опухоли RNF43 представляет собой стволовую клетку Лигаза Е3, индуцирующая эндоцитоз рецепторов Wnt. Природа. 2012, 488 (7413): 665-669. 10.1038/nature11308
CAS пабмед Статья Google Scholar
Хао Х.С., Се И., Чжан И., Шарлат О., Остер Э., Авелло М., Лей Х., Миканин С., Лю Д., Руффнер Х., Мао Х., Ма К., Дзампони Р., Боумистер Т., Финан П.М., Киршнер М.В., Портер JA, Serluca FC, Cong F: ZNRF3 способствует обороту рецептора Wnt чувствительным к R-спондину образом. Природа. 2012, 485 (7397): 195-200. 10.1038/nature11019
CAS пабмед Статья Google Scholar

Mukai A, Yamamoto-Hino M, Awano W, Watanabe W, Komada M, Goto S: Сбалансированное убиквитинирование и деубиквитилирование Frizzled регулирует клеточную реакцию на Wg/Wnt.EMBO J. 2010, 29 (13): 2114-2125. 10.1038/emboj.2010.100
PubMed Central КАС пабмед Статья Google Scholar
Yamamoto A, Nagano T, Takehara S, Hibi M, Aizawa S: Shisa способствует формированию головок за счет ингибирования созревания рецепторных белков для каудальных факторов, Wnt и FGF. Клетка. 2005, 120 (2): 223-235. 10.1016/j.cell.2004.11.051
CAS пабмед Статья Google Scholar
Davidson G, Wu W, Shen J, Bilic J, Fenger U, Stannek P, Glinka A, Niehrs C: Казеинкиназа 1γ связывает активацию рецептора Wnt с передачей цитоплазматического сигнала. Природа. 2005, 438 (7069): 867-872. 10.1038/nature04170
CAS пабмед Статья Google Scholar

Zeng X, Tamai K, Doble B, Li S, Huang H, Habas R, Okamura H, Woodgett J, He X: Механизм двойной киназы для фосфорилирования и активации корецептора Wnt.Природа. 2005, 438 (7069): 873-877. 10.1038/nature04185
PubMed Central КАС пабмед Статья Google Scholar
MacDonald BT, Yokota C, Tamai K, Zeng X, He X: усиление сигнала Wnt за счет активности, кооперативности и регуляции множественных внутриклеточных мотивов PPPSP в корецепторе Wnt LRP6. Дж. Биол. Хим. 2008, 283 (23): 16115-16123. 10.1074/jbc.M800327200
Центральный пабмед КАС пабмед Статья Google Scholar
Chen M, Philipp M, Wang J, Premont RT, Garrison TR, Caron MG, Lefkowitz RJ, Chen W: Киназы рецептора, связанного с G-белком, фосфорилируют LRP6 в пути Wnt. Дж. Биол. Хим. 2009, 284 (50): 35040-35048. 10.1074/jbc.M109.047456
PubMed Central КАС пабмед Статья Google Scholar

Червенка И., Вольф Дж., Машек Дж., Крейци П., Уилкокс В.Р., Козубик А., Шульте Г., Гуткинд Дж.С., Брия В.: Митоген-активируемые протеинкиназы способствуют передаче сигналов WNT/β-катенина посредством фосфорилирования LRP6.Мол Селл Биол. 2011, 31 (1): 179-189. 10.1128/MCB.00550-10
Центральный пабмед пабмед Статья КАС Google Scholar
Святек В., Канг Х., Гарсия Б.А., Шабановиц Дж., Кумбс Г.С., Хант Д.Ф., Виршуп Д.М.: Негативная регуляция функции LRP6 фосфорилированием казеинкиназы Iϵ. Дж. Биол. Хим. 2006, 281 (18): 12233-12241. 10.1074/джбк.М510580200
КАС пабмед Статья Google Scholar
Wan M, Yang C, Li J, Wu X, Yuan H, Ma H, He X, Nie S, Chang C, Cao X: передача сигналов паратиреоидного гормона через белок, родственный липопротеинам низкой плотности 6. Genes Dev. 2008, 22 (21): 2968-2979. 10.1101/гад.1702708
PubMed Central КАС пабмед Статья Google Scholar

Davidson G, Shen J, Huang YL, Su Y, Karaulanov E, Bartscherer K, Hassler C, Stannek P, Boutros M, Niehrs C: Контроль клеточного цикла активации рецептора Wnt.Ячейка Дев. 2009, 17 (6): 788-799. 10.1016/j.devcel.2009.11.006
CAS пабмед Статья Google Scholar
Abrami L, Kunz B, Iacovache I, van der Goot FG: Пальмитоилирование и убиквитинирование регулируют выход сигнального белка Wnt LRP6 из эндоплазматического ретикулума. Proc Natl Acad Sci USA. 2008, 105 (14): 5384-5389. 10.1073/pnas.0710389105
PubMed Central КАС пабмед Статья Google Scholar
Макдональд Б.Т., Семенов М.В., Хуан Х., Хе Х. Анализ молекулярных различий между корецепторами Wnt LRP5 и LRP6. ПЛОС Один. 2011, 6 (8): e23537. 10.1371/journal.pone.0023537
PubMed Central КАС пабмед Статья Google Scholar

Ikeda S, Kishida S, Yamamoto H, Murai H, Koyama S, Kikuchi A: Axin, негативный регулятор сигнального пути Wnt, образует комплекс с GSK-3β и β-катенином и способствует GSK- 3β-зависимое фосфорилирование β-катенина.EMBO J. 1998, 17 (5): 1371-1384. 10.1093/emboj/17.5.1371
Центральный пабмед КАС пабмед Статья Google Scholar
Yamamoto H, Kishida S, Kishida M, Ikeda S, Takada S, Kikuchi A: Фосфорилирование аксина, отрицательного регулятора сигнала Wnt, киназой-3β гликогенсинтазы регулирует его стабильность. Дж. Биол. Хим. 1999, 274 (16): 10681-10684. 10.1074/джбк.274.16.10681
КАС пабмед Статья Google Scholar
Jho E, Lomvardas S, Costantini F: Сайт фосфорилирования GSK3β в аксине модулирует взаимодействие с β-катенином и Tcf-опосредованную экспрессию генов. Biochem Biophys Res Commun. 1999, 266 (1): 28-35. 10.1006/bbrc.1999. 1760
1760
CAS пабмед Статья Google Scholar
Чжан Ю., Лю С., Миканин С., Фэн Ю., Шарла О., Мишо Г.А., Ширле М., Ши Х., Хильд М., Бауэр А., Майер В.Е., Финан П.М., Портер Дж.А., Хуан С.М., Конг Ф. : RNF146 представляет собой поли(АДФ-рибозо)-направленную лигазу E3, которая регулирует деградацию аксина и передачу сигналов Wnt.Nat Cell Biol. 2011, 13 (5): 623-629. 10.1038/ncb2222
КАС пабмед Статья Google Scholar
Кэллоу М.Г., Тран Х., Фу Л., Лау Т., Ли Дж., Сандовал В.Н., Лю П.С., Бхеддах С., Тао Дж., Лилль Дж.Р., Хонго Дж.А., Дэвис Д., Киркпатрик Д.С., Полакис П., Коста М. : Убиквитинлигаза RNF146 регулирует танкиразу и аксин, чтобы способствовать передаче сигналов Wnt. ПЛОС Один. 2011, 6 (7): e22595. 10.1371/journal.pone.0022595
PubMed Central КАС пабмед Статья Google Scholar

Fei C, Li Z, Li C, Chen Y, Chen Z, He X, Mao L, Wang X, Zeng R, Li L: Smurf1-опосредованное Lys29-связанное полиубиквитинирование Axin отрицательно регулирует передачу сигналов Wnt/β-катенина. Мол Селл Биол. 2013, 33 (20): 4095-4105. 10.1128/MCB.00418-13
Центр ПабМед КАС пабмед Статья Google Scholar
Kim S, Jho EH: Стабильность белка Axin, отрицательного регулятора передачи сигналов Wnt, регулируется фактором 2 регуляции убиквитинирования Smad (Smurf2).Дж. Биол. Хим. 2010, 285 (47): 36420-36426. 10.1074/jbc.M110.137471
Центральный пабмед КАС пабмед Статья Google Scholar
Rui HL, Fan E, Zhou H-M, Xu Z, Zhang Y, Lin SC: модификация SUMO-1 C-терминала KVEKVD Axin необходима для активации JNK, но не влияет на передачу сигналов Wnt. Дж. Биол. Хим. 2002, 277 (45): 42981-42986. 10.1074/джбк.М208099200
КАС пабмед Статья Google Scholar

Хуан С.М., Мишина Ю.М., Лю С., Чунг А., Стегмайер Ф., Мишо Г.А., Шарла О., Виеллетт Э., Чжан Ю., Висснер С., Хильд М., Ши Х., Уилсон С.Дж., Миканин С., Майер В., Фазал А., Томлинсон R, Serluca F, Shao W, Cheng H, Shultz M, Rau C, Schirle M, Schlegl J, Ghidelli S, Fawell S, Lu C, Curtis D, Kirschner MW, Lengauer C: Ингибирование танкиразы стабилизирует аксин и противодействует передаче сигналов Wnt. Природа. 2009, 461 (7264): 614-620. 10.1038/nature08356
CAS пабмед Статья Google Scholar
Кросс Д.А., Алесси Д.Р., Коэн П., Анджелкович М., Хеммингс Б.А.: Ингибирование киназы-3 гликогенсинтазы инсулином, опосредованное протеинкиназой B. Nature. 1995, 378 (6559): 785-789. 10.1038/378785a0
КАС пабмед Статья Google Scholar
Fang X, Yu SX, Lu Y, Bast RC, Woodgett JR, Mills GB: Фосфорилирование и инактивация киназы гликогенсинтазы 3 протеинкиназой A. Proc Natl Acad Sci US A. 2000, 97 (22) : 11960-11965.10.1073/pnas.220413597
Proc Natl Acad Sci US A. 2000, 97 (22) : 11960-11965.10.1073/pnas.220413597
PubMed Central КАС пабмед Статья Google Scholar
Ballou LM, Tian PY, Lin HY, Jiang YP, Lin RZ: Двойная регуляция киназы-3β гликогенсинтазы с помощью α1A-адренергического рецептора. Дж. Биол. Хим. 2001, 276 (44): 40910-40916. 10.1074/джбк.М103480200
КАС пабмед Статья Google Scholar
Doble BW, Woodgett JR: GSK-3: хитрости для многозадачной киназы.Дж. Клеточные науки. 2003, 116 (7): 1175-1186. 10.1242/jcs.00384
Центральный пабмед КАС пабмед Статья Google Scholar
Zhang HH, Lipovsky AI, Dibble CC, Sahin M, Manning BD: S6K1 регулирует GSK3 в условиях mTOR-зависимого ингибирования Akt. Мол Ячейка. 2006, 24 (2): 185-197. 10.1016/j.molcel.2006.09.019
PubMed Central КАС пабмед Статья Google Scholar

Hughes K, Nikolakaki E, Plyte SE, Totty NF, Woodgett JR: Модуляция семейства киназ-3 гликогенсинтазы путем фосфорилирования тирозина. EMBO J. 1993, 12 (2): 803-808.
Центральный пабмед КАС пабмед Google Scholar
Sayas CL, Ariaens A, Ponsioen B, Moolenaar WH: GSK-3 активируется тирозинкиназой Pyk2 во время LPA1-опосредованной ретракции нейритов. Мол Биол Селл. 2006, 17 (4): 1834-1844. 10.1091/мбк.E05-07-0688
Центральный пабмед КАС пабмед Статья Google Scholar
Cole A, Frame S, Cohen P: Еще одно доказательство того, что фосфорилирование тирозина киназы-3 гликогенсинтазы (GSK3) в клетках млекопитающих является событием аутофосфорилирования. Biochem J. 2004, 377 (часть 1): 249-255.
Центральный пабмед КАС пабмед Статья Google Scholar
Ding Q, Xia W, Liu JC, Yang JY, Lee DF, Xia J, Bartholomeusz G, Li Y, Pan Y, Li Z, Bargou RC, Qin J, Lai CC, Tsai FJ, Tsai CH, Hung MC: Erk связывается с GSK-3β и инициирует его инактивацию, что приводит к усилению регуляции β-катенина. Мол Ячейка. 2005, 19 (2): 159-170. 10.1016/j.molcel.2005.06.009
Мол Ячейка. 2005, 19 (2): 159-170. 10.1016/j.molcel.2005.06.009
CAS пабмед Статья Google Scholar
Thornton TM, Pedraza-Alva G, Deng B, Wood CD, Aronshtam A, Clements JL, Sabio G, Davis RJ, Matthews DE, Doble B, Rincon M: Фосфорилирование p38 MAPK как альтернативный путь для Инактивация GSK3β.Наука. 2008, 320 (5876): 667-670. 10.1126/science.1156037
PubMed Central КАС пабмед Статья Google Scholar
Файлор К.Л., Десятников Ю., Фингер Л.А., Файрстоун Г.Л.: Глюкокортикоид-индуцированная деградация белка гликогенсинтаза-киназы-3 запускается сывороточной и глюкокортикоид-индуцированной протеинкиназой и передачей сигналов Akt и контролирует динамику β-катенина и образование плотных контактов в эпителиальных опухолевых клетках молочной железы.Мол Эндокринол. 2007, 21 (10): 2403-2415. 10.1210/ме.2007-0143
КАС пабмед Статья Google Scholar

Eun Jeoung L, Sung Hee H, Jaesun C, Sung Hwa S, Kwang Hum Y, Min Kyoung K, Tae Yoon P, Sang Sun K: Регуляция функций киназы гликогенсинтазы 3beta путем модификации малого убиквитина -подобный модификатор. Open Biochem J. 2008, 2: 67-76.
ПабМед Статья КАС Google Scholar
Feijs KL, Kleine H, Braczynski A, Forst A, Herzog N, Verheugd P, Linzen U, Kremmer E, Luscher B: Идентификация субстрата ARTD10 на белковых микрочипах: регуляция GSK3beta путем моно-АДФ-рибозилирования. Сигнал сотовой связи. 2013, 11 (1): 5. 10.1186/1478-811X-11-5
PubMed Central КАС пабмед Статья Google Scholar
Ikeda S, Kishida M, Matsuura Y, Usui H, Kikuchi A: GSK-3β-зависимое фосфорилирование продукта гена аденоматозного полипоза кишечной палочки может модулироваться бета-катенином и протеинфосфатазой 2A в комплексе с аксином.Онкоген. 2000, 19 (4): 537-545. 10.1038/sj.onc.1203359
10.1038/sj.onc.1203359
CAS пабмед Статья Google Scholar
Rubinfeld B, Albert I, Porfiri E, Fiol C, Munemitsu S, Polakis P: Связывание GSK3β с комплексом APC-β-катенин и регуляция сборки комплекса. Наука. 1996, 272 (5264): 1023-1026. 10.1126/наука.272.5264.1023
CAS пабмед Статья Google Scholar
Ha NC, Tonozuka T, Stamos JL, Choi HJ, Weis WI: Механизм зависимого от фосфорилирования связывания APC с β-катенином и его роль в деградации β-катенина. Мол Ячейка. 2004, 15 (4): 511-521. 10.1016/j.molcel.2004.08.010
CAS пабмед Статья Google Scholar
Рубинфельд Б., Тайс Д.А., Полакис П.: Аксинозависимое фосфорилирование белка аденоматозного полипоза толстой кишки, опосредованное казеинкиназой 1ϵ. Дж. Биол. Хим.2001, 276 (42): 39037-39045. 10.1074/джбк.М105148200
КАС пабмед Статья Google Scholar

Choi J, Park SY, Costantini F, Jho E-h, Joo C-K: Аденоматозный полипоз толстой кишки подавляется убиквитин-протеасомным путем в процессе, которому способствует Axin. Дж. Биол. Хим. 2004, 279 (47): 49188-49198. 10.1074/джбк.М404655200
КАС пабмед Статья Google Scholar
Huang X, Langelotz C, Hetfeld-Pěchoč BK, Schwenk W, Dubiel W: Сигналосома COP9 опосредует деградацию β-катенина путем денеддиляции и блокирует разрушение аденоматозного полипоза толстой кишки через USP15. Дж Мол Биол. 2009, 391 (4): 691-702. 10.1016/j.jmb.2009.06.066
КАС пабмед Статья Google Scholar
Tran H, Hamada F, Schwarz-Romond T, Bienz M: Trabid, новый положительный регулятор Wnt-индуцированной транскрипции с предпочтением связывания и расщепления K63-связанных цепей убиквитина.Гены Дев. 2008, 22 (4): 528-542. 10.1101/гад.463208
PubMed Central КАС пабмед Статья Google Scholar

Tran H, Bustos D, Yeh R, Rubinfeld B, Lam C, Shriver S, Zilberleyb I, Lee MW, Phu L, Sarkar AA, Zohn IE, Wertz IE, Kirkpatrick DS, Polakis P: HectD1 E3 лигаза модифицирует аденоматозный полипоз толстой кишки (APC) с помощью полиубиквитина, чтобы способствовать взаимодействию APC-Axin. Дж. Биол. Хим. 2013, 288 (6): 3753-3767. 10.1074/jbc.M112.415240
Центральный пабмед КАС пабмед Статья Google Scholar
Климовски Л.К., Гарсия Б.А., Шабановиц Дж., Хант Д.Ф., Виршуп Д.М.: Сайт-специфическое казеинкиназное 1ϵ-зависимое фосфорилирование Disheveled модулирует передачу сигналов β-катенина. FEBS J. 2006, 273 (20): 4594-4602. 10.1111/j.1742-4658.2006.05462.x
CAS пабмед Статья Google Scholar
Hino S, Michiue T, Asashima M, Kikuchi A: Казеинкиназа Iϵ усиливает связывание Dvl-1 с Frat-1 и необходима для индуцированного Wnt-3a накопления β-катенина. Дж. Биол. Хим. 2003, 278 (16): 14066-14073. 10.1074/джбк.М213265200
Дж. Биол. Хим. 2003, 278 (16): 14066-14073. 10.1074/джбк.М213265200
КАС пабмед Статья Google Scholar
Хуан Х., МакГанн Дж. К., Лю Б.Я., Ханнуш Р.Н., Лилл Дж. Р., Фам В., Ньютон К., Какунда М., Лю Дж., Ю. С., Химовиц С. Г., Хонго Дж. А., Уиншоу-Борис А., Полакис П., Harland RM, Dixit VM: Фосфорилирование взлохмаченных протеинкиназой RIPK4 регулирует передачу сигналов Wnt.Наука. 2013, 339 (6126): 1441-1445. 10.1126/science.1232253
PubMed Central КАС пабмед Статья Google Scholar
Klein TJ, Jenny A, Djiane A, Mlodzik M: Заросшие CKIε/диски способствуют передаче сигналов Wnt-Fz/β-catenin и Fz/PCP у дрозофилы. Карр Биол. 2006, 16 (13): 1337-1343. 10.1016/j.cub.2006.06.030
CAS пабмед Статья Google Scholar
Sun TQ, Lu B, Feng JJ, Reinhard C, Jan YN, Fantl WJ, Williams LT: PAR-1 представляет собой киназу, связанную с Dishavelled, и позитивный регулятор передачи сигналов Wnt. Nat Cell Biol. 2001, 3 (7): 628-636. 10.1038/35083016
Nat Cell Biol. 2001, 3 (7): 628-636. 10.1038/35083016
КАС пабмед Статья Google Scholar
Willert K, Brink M, Wodarz A, Varmus H, Nusse R: Казеинкиназа 2 связывается с Dishavelled и фосфорилирует его. EMBO J. 1997, 16 (11): 3089-3096. 10.1093/emboj/16.11.3089
Центральный пабмед КАС пабмед Статья Google Scholar
Cruciat CM, Dolde C, de Groot RE, Ohkawara B, Reinhard C, Korswagen HC, Niehrs C: РНК-хеликаза DDX3 является регуляторной субъединицей казеинкиназы 1 в передаче сигналов Wnt-β-катенин. Наука. 2013, 339 (6126): 1436-1441. 10.1126/наука.1231499
CAS пабмед Статья Google Scholar
Jung H, Kim BG, Han WH, Lee JH, Cho JY, Park WS, Maurice MM, Han JK, Lee MJ, Finley D, Jho EH: для передачи сигналов Wnt требуется деубиквитинирование Disheveled с помощью Usp14. Онкогенез. 2013, 2: е64. 10.1038/oncsis.2013.28
Онкогенез. 2013, 2: е64. 10.1038/oncsis.2013.28
PubMed Central КАС пабмед Статья Google Scholar
Tauriello DV, Haegebarth A, Kuper I, Edelmann MJ, Henraat M, Canninga-van Dijk MR, Kessler BM, Clevers H, Maurice MM: Потеря опухолевого супрессора CYLD усиливает передачу сигналов Wnt/β-катенина посредством K63-сцепленное убиквитинирование Dvl.Мол Ячейка. 2010, 37 (5): 607-619. 10.1016/j.molcel.2010.01.035
CAS пабмед Статья Google Scholar
Angers S, Thorpe CJ, Biechele TL, Goldenberg SJ, Zheng N, MacCoss MJ, Moon RT: Убиквитинлигаза KLHL12-Cullin-3 негативно регулирует путь Wnt-[beta]-катенина, воздействуя на Disheveled для деградация. Nat Cell Biol. 2006, 8 (4): 348-357. 10.1038/нкб1381
КАС пабмед Статья Google Scholar
Wei W, Li M, Wang J, Nie F, Li L: Убиквитинлигаза E3 ITCH негативно регулирует каноническую передачу сигналов Wnt, нацеливаясь на растрепанный белок. Мол Селл Биол. 2012, 32 (19): 3903-3912. 10.1128/MCB.00251-12
Мол Селл Биол. 2012, 32 (19): 3903-3912. 10.1128/MCB.00251-12
Центральный пабмед КАС пабмед Статья Google Scholar
Ding Y, Zhang Y, Xu C, Tao QH, Chen YG: убиквитинлигаза E3, содержащая домен HECT, NEDD4L отрицательно регулирует передачу сигналов Wnt, нацеливая растрепанные на протеасомную деградацию.Дж. Биол. Хим. 2013, 288 (12): 8289-8298. 10.1074/jbc.M112.433185
Центральный пабмед КАС пабмед Статья Google Scholar
Gao C, Cao W, Bao L, Zuo W, Xie G, Cai T, Fu W, Zhang J, Wu W, Zhang X, Chen YG: Аутофагия негативно регулирует передачу сигналов Wnt, способствуя деградации Disheveled. Nat Cell Biol. 2010, 12 (8): 781-790. 10.1038/ncb2082
КАС пабмед Статья Google Scholar
Шарма Дж., Малхеркар С., Мукерджи Д., Яна Н.Р.: Малин регулирует сигнальный путь Wnt посредством деградации растрепанный2. Дж. Биол. Хим. 2012, 287 (9): 6830-6839. 10.1074/jbc.M111.315135
Дж. Биол. Хим. 2012, 287 (9): 6830-6839. 10.1074/jbc.M111.315135
Центральный пабмед КАС пабмед Статья Google Scholar
Miyazaki K, Fujita T, Ozaki T, Kato C, Kurose Y, Sakamoto M, Kato S, Goto T, Itoyama Y, Aoki M, Nakagawara A: NEDL1, новая убиквитин-протеинизопептидная лигаза для взъерошенных -1, нацеливается на мутантную супероксиддисмутазу-1.Дж. Биол. Хим. 2004, 279 (12): 11327-11335. 10.1074/джбк.М312389200
КАС пабмед Статья Google Scholar
Yost C, Torres M, Miller JR, Huang E, Kimelman D, Moon RT: индуцирующая ось активность, стабильность и субклеточное распределение бета-катенина регулируются у эмбрионов Xenopus киназой гликогенсинтазы 3. Гены Дев. 1996, 10 (12): 1443-1454. 10.1101/гад.10.12.1443
КАС пабмед Статья Google Scholar
Пайфер М. , Пай Л.М., Кейси М.: Фосфорилирование соединительного белка дрозофилы Armadillo: роль бескрылого сигнала и киназы zeste-white 3. Дев биол. 1994, 166 (2): 543-556. 10.1006/dbio.1994.1336
, Пай Л.М., Кейси М.: Фосфорилирование соединительного белка дрозофилы Armadillo: роль бескрылого сигнала и киназы zeste-white 3. Дев биол. 1994, 166 (2): 543-556. 10.1006/dbio.1994.1336
CAS пабмед Статья Google Scholar
Hino S, Tanji C, Nakayama KI, Kikuchi A: Фосфорилирование β-катенина циклической АМФ-зависимой протеинкиназой стабилизирует β-катенин за счет ингибирования его убиквитинирования.Мол Селл Биол. 2005, 25 (20): 9063-9072. 10.1128/MCB.25.20.9063-9072.2005
PubMed Central КАС пабмед Статья Google Scholar
Fang D, Hawke D, Zheng Y, Xia Y, Meisenhelder J, Nika H, Mills GB, Kobayashi R, Hunter T, Lu Z: Фосфорилирование β-катенина с помощью AKT способствует транскрипционной активности β-катенина. Дж. Биол. Хим. 2007, 282 (15): 11221-11229. 10.1074/jbc.M611871200
Центральный пабмед КАС пабмед Статья Google Scholar

Wu X, Tu X, Joeng KS, Hilton MJ, Williams DA, Long F: активация Rac1 контролирует ядерную локализацию β-катенина во время канонической передачи сигналов Wnt. Клетка. 2008, 133 (2): 340-353. 10.1016/j.cell.2008.01.052
Центральный пабмед КАС пабмед Статья Google Scholar
Du C, Zhang C, Li Z, Biswas MH, Balaji KC: β-катенин, фосфорилированный по треонину 120, препятствует образованию активного β-катенина за счет пространственной локализации в сети транс-Гольджи.ПЛОС Один. 2012, 7 (4): e33830. 10.1371/journal.pone.0033830
PubMed Central КАС пабмед Статья Google Scholar
Wu G, Xu G, Schulman BA, Jeffrey PD, Harper JW, Pavletich NP: Структура комплекса β-TrCP1-Skp1-β-катенин: связывание деструктивного мотива и лизиновая специфичность SCF β- TrCP1 убиквитинлигаза. Мол Ячейка. 2003, 11 (6): 1445-1456. 10.1016/С1097-2765(03)00234-С
КАС пабмед Статья Google Scholar

Winer IS, Bommer GT, Gonik N, Fearon ER: Лизиновые остатки Lys-19 и Lys-49 β-катенина регулируют его уровни и функцию в активации транскрипции Т-клеточного фактора и неопластической трансформации. Дж. Биол. Хим. 2006, 281 (36): 26181-26187. 10.1074/джбк.М604217200
КАС пабмед Статья Google Scholar
Китагава М., Хатакеяма С., Ширане М., Мацумото М., Исида Н., Хаттори К., Накамичи И., Кикучи А., Накаяма К., Накаяма К.: Белок F-бокса, FWD1, опосредует убиквитин-зависимый протеолиз β-катенин.EMBO J. 1999, 18 (9): 2401-2410. 10.1093/emboj/18.9.2401
Центральный пабмед КАС пабмед Статья Google Scholar
Latres E, Chiaur DS, Pagano M: Белок F-бокса человека β-Trcp связывается с комплексом Cul1/Skp1 и регулирует стабильность бета-катенина. Онкоген. 1999, 18 (4): 849-854. 10.1038/sj.onc.1202653
CAS пабмед Статья Google Scholar

Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, Harper JW: Комплекс SCF β-TRCP -убиквитинлигаза специфически связывается с мотивами фосфорилированного разрушения в IκBα и β-катенине и стимулирует убиквитинирование IκBα в витро . Гены Дев. 1999, 13 (3): 270-283. 10.1101/gad.13.3.270
PubMed Central КАС пабмед Статья Google Scholar
Hay-Koren A, Caspi M, Zilberberg A, Rosin-Arbesfeld R: Убиквитинлигаза EDD E3 убиквитинирует и активирует β-катенин.Мол Биол Селл. 2011, 22 (3): 399-411. 10.1091/mbc.E10-05-0440
Центр PubMed КАС пабмед Статья Google Scholar
Shekhar MP, Gerard B, Pauley RJ, Williams BO, Tait L: Rad6B является положительным регулятором стабилизации β-катенина. Рак рез. 2008, 68 (6): 1741-1750. 10.1158/0008-5472.КАН-07-2111
КАС пабмед Статья Google Scholar

Gerard B, Sanders MA, Visscher DW, Tait L, Shekhar MP: Лизин 394 представляет собой новый Rad6B-индуцированный сайт убиквитинирования на β-катенине. Биохим Биофиз Акта. 2012, 1823 (10): 1686-1696. 10.1016/j.bbamcr.2012.05.032
PubMed Central КАС пабмед Статья Google Scholar
Dao KH, Rotelli MD, Petersen CL, Kaech S, Nelson WD, Yates JE, Hanlon Newell AE, Olson SB, Druker BJ, Bagby GC: FANCL убиквитинирует β-катенин и усиливает его ядерную функцию.Кровь. 2012, 120 (2): 323-334. 10.1182/blood-2011-11-388355
PubMed Central КАС пабмед Статья Google Scholar
Chitalia VC, Foy RL, Bachschmid MM, Zeng L, Panchenko MV, Zhou MI, Bharti A, Seldin DC, Lecker SH, Dominguez I, Cohen HT: Jade-1 ингибирует передачу сигналов Wnt путем убиквитилирования β-катенина и опосредует ингибирование пути Wnt с помощью pVHL. Nat Cell Biol. 2008, 10 (10): 1208-1216. 10.1038/ncb1781
10.1038/ncb1781
Центральный пабмед КАС пабмед Статья Google Scholar
Wolf D, Rodova M, Miska EA, Calvet JP, Kouzarides T: Ацетилирование β-катенина CREB-связывающим белком (CBP). Дж. Биол. Хим. 2002, 277 (28): 25562-25567. 10.1074/джбк.М201196200
КАС пабмед Статья Google Scholar
Lévy L, Wei Y, Labalette C, Wu Y, Renard CA, Buendia MA, Neuveut C: Ацетилирование β-катенина с помощью p300 регулирует взаимодействие β-катенин-Tcf4. Мол Селл Биол. 2004, 24 (8): 3404-3414. 10.1128/МКБ.24.8.3404-3414.2004
Центральный пабмед пабмед Статья КАС Google Scholar
Ge X, Jin Q, Zhang F, Yan T, Zhai Q: PCAF ацетилирует β-катенин и повышает его стабильность. Мол Биол Селл. 2009, 20 (1): 419-427. 10.1091/mbc.E08-08-0792
PubMed Central КАС пабмед Статья Google Scholar

Ishitani T, Ninomiya-Tsuji J, Matsumoto K: Регуляция лимфоидного энхансерного фактора 1/T-клеточного фактора с помощью митоген-активируемой протеинкиназы, связанной с Nemo-like kinase-зависимым фосфорилированием в передаче сигналов Wnt/β-catenin .Мол Селл Биол. 2003, 23 (4): 1379-1389. 10.1128/MCB.23.4.1379-1389.2003
PubMed Central КАС пабмед Статья Google Scholar
Ишитани Т., Ниномия-Цудзи Дж., Нагаи С., Нишита М., Менегини М., Баркер Н., Уотерман М., Бауэрман Б., Клеверс Х., Сибуя Х., Мацумото К. Путь, связанный с TAK1-NLK-MAPK противодействует передаче сигналов между β-катенином и фактором транскрипции TCF. Природа. 1999, 399 (6738): 798-802. 10.1038/21674
КАС пабмед Статья Google Scholar
Mahmoudi T, Li VSW, Ng SS, Taouatas N, Vries RGJ, Mohammed S, Heck AJ, Clevers H: киназа TNIK является важным активатором генов-мишеней Wnt. EMBO J. 2009, 28 (21): 3329-3340. 10.1038/emboj.2009.285
EMBO J. 2009, 28 (21): 3329-3340. 10.1038/emboj.2009.285
PubMed Central КАС пабмед Статья Google Scholar
Shitashige M, Satow R, Jigami T, Aoki K, Honda K, Shibata T, Ono M, Hirohashi S, Yamada T: киназа, взаимодействующая с Traf2 и Nck, необходима для передачи сигналов Wnt и роста колоректального рака.Рак рез. 2010, 70 (12): 5024-5033. 10.1158/0008-5472.КАН-10-0306
КАС пабмед Статья Google Scholar
Lee E, Salic A, Kirschner MW: Физиологическая регуляция стабильности β-катенина с помощью Tcf3 и CK1∈. Джей Селл Биол. 2001, 154 (5): 983-994. 10.1083/jcb.200102074
Центр PubMed КАС пабмед Статья Google Scholar
Wang S, Jones KA: CK2 контролирует привлечение регуляторов Wnt к генам-мишеням in vivo. Карр Биол. 2006, 16 (22): 2239-2244. 10.1016/j. cub.2006.09.034
cub.2006.09.034
CAS пабмед Статья Google Scholar
Hämmerlein A, Weiske J, Huber O: Вторая стадия, опосредованная протеинкиназой CK1, негативно регулирует передачу сигналов Wnt, разрушая комплекс энхансера лимфоцитов фактор-1/β-катенин. Cell Mol Life Sci. 2005, 62 (5): 606-618.10.1007/s00018-005-4507-7
PubMed Статья КАС Google Scholar
Hikasa H, Sokol SY: Фосфорилирование белков TCF протеинкиназой, взаимодействующей с гомеодоменом 2. J Biol Chem. 2011, 286 (14): 12093-12100. 10.1074/jbc.M110.185280
Центральный пабмед КАС пабмед Статья Google Scholar
Hikasa H, Ezan J, Itoh K, Li X, Klymkowsky MW, Sokol SY: Регуляция TCF3 Wnt-зависимым фосфорилированием во время спецификации оси позвоночных.Ячейка Дев. 2010, 19 (4): 521-532. 10.1016/j.devcel. 2010.09.005
2010.09.005
Центральный пабмед КАС пабмед Статья Google Scholar
Waltzer L, Bienz M: Drosophila CBP репрессирует фактор транскрипции TCF, чтобы противодействовать передаче сигналов Wingless. Природа. 1998, 395 (6701): 521-525. 10.1038/26785
КАС пабмед Статья Google Scholar
Gay F, Calvo D, Lo MC, Ceron J, Maduro M, Lin R, Shi Y: Ацетилирование регулирует субклеточную локализацию сигнального ядерного эффектора Wnt POP-1.Гены Дев. 2003, 17 (6): 717-722. 10.1101/гад.1042403
PubMed Central КАС пабмед Статья Google Scholar
Elfert S, Weise A, Bruser K, Biniossek ML, Jägle S, Senghaas N, Hecht A: Ацетилирование белков TCF4 (TCF7L2) человека ослабляет ингибирование репрессором HBP1 и вызывает конформационные изменения в TCF4: : Комплекс ДНК. ПЛОС Один. 2013, 8 (4): e61867. 10.1371/journal.pone.0061867
ПЛОС Один. 2013, 8 (4): e61867. 10.1371/journal.pone.0061867
PubMed Central КАС пабмед Статья Google Scholar
Sachdev S, Bruhn L, Sieber H, Pichler A, Melchior F, Grosschedl R: PIASy, связанная с ядерным матриксом лигаза SUMO E3, подавляет активность LEF1 путем секвестрации в ядерные тельца. Гены Дев. 2001, 15 (23): 3088-3103. 10.1101/гад.944801
PubMed Central КАС пабмед Статья Google Scholar
Yamamoto H, Ihara M, Matsuura Y, Kikuchi A: Сумоилирование участвует в β-катенин-зависимой активации Tcf-4.EMBO J. 2003, 22 (9): 2047-2059. 10.1093/emboj/cdg204
Центральный PubMed КАС пабмед Статья Google Scholar
Ishitani T, Matsumoto K, Chitnis AB, Itoh M: функции Nrarp модулируют дифференцировку клеток нервного гребня, регулируя стабильность белка LEF1. Nat Cell Biol. 2005, 7 (11): 1106-1112. 10.1038/ncb1311
Nat Cell Biol. 2005, 7 (11): 1106-1112. 10.1038/ncb1311
КАС пабмед Статья Google Scholar
Yamada M, Ohnishi J, Ohkawara B, Iemura S, Satoh K, Hyodo-Miura J, Kawachi K, Natsume T, Shibuya H: NARF, немоподобная киназа (NLK), ассоциированный с безымянным пальцем белок, регулирует убиквитилирование и деградацию фактора Т-клеток/лимфоидного энхансера (TCF/LEF). Дж. Биол. Хим. 2006, 281 (30): 20749-20760. 10.1074/джбк.М602089200
КАС пабмед Статья Google Scholar
Коэн П. Роль фосфорилирования белка в здоровье и болезни человека.Евр Дж Биохим. 2001, 268 (19): 5001-5010. 10.1046/j.0014-2956.2001.02473.x
CAS пабмед Статья Google Scholar
Aberle H, Bauer A, Stappert J, Kispert A, Kemler R: β-катенин является мишенью убиквитин-протеасомного пути. EMBO J. 1997, 16 (13): 3797-3804. 10.1093/emboj/16.13.3797
1997, 16 (13): 3797-3804. 10.1093/emboj/16.13.3797
PubMed Central КАС пабмед Статья Google Scholar
Hart M, Concordet JP, Lassot I, Albert I, del los Santos R, Durand H, Perret C, Rubinfeld B, Margottin F, Benarous R, Polakis P: Белок F-бокса β-TrCP связывается с фосфорилированным β-катенином и регулирует его активность в клетке. Карр Биол. 1999, 9 (4): 207-211. 10.1016/С0960-9822(99)80091-8
КАС пабмед Статья Google Scholar
Liu X, Rubin JS, Kimmel AR: Быстрые, индуцированные Wnt изменения в ассоциациях GSK3β, которые регулируют стабилизацию β-катенина, опосредуются белками Gα.Карр Биол. 2005, 15 (22): 1989-1997. 10.1016/j.cub.2005.10.050
CAS пабмед Статья Google Scholar
Cselenyi CS, Jernigan KK, Tahinci E, Thorne CA, Lee LA, Lee E: LRP6 передает канонический сигнал Wnt независимо от деградации Axin путем ингибирования фосфорилирования GSK3 β-катенина. Proc Natl Acad Sci USA. 2008, 105 (23): 8032-8037. 10.1073/pnas.0803025105
Proc Natl Acad Sci USA. 2008, 105 (23): 8032-8037. 10.1073/pnas.0803025105
PubMed Central КАС пабмед Статья Google Scholar
Piao S, Lee SH, Kim H, Yum S, Stamos JL, Xu Y, Lee SJ, Lee J, Oh S, Han JK, Park BJ, Weis WI, Ha NC: Прямое ингибирование GSK3β фосфорилированным цитоплазматическим доменом LRP6 в передаче сигналов Wnt/β-катенина. ПЛОС Один. 2008, 3 (12): e4046. 10.1371/journal.pone.0004046
PubMed Central пабмед Статья КАС Google Scholar
Wu G, Huang H, Abreu JG, He X: Ингибирование GSK3 фосфорилирования β-катенина посредством фосфорилированных мотивов PPPSPXS корецептора Wnt LRP6.ПЛОС Один. 2009, 4 (3): e4926. 10.1371/journal.pone.0004926
PubMed Central пабмед Статья КАС Google Scholar
Mi K, Dolan PJ, Johnson GVW: Белок 6, родственный рецептору липопротеинов низкой плотности, взаимодействует с киназой гликогенсинтазы 3 и ослабляет активность. Дж. Биол. Хим. 2006, 281 (8): 4787-4794. 10.1074/джбк.М508657200
Дж. Биол. Хим. 2006, 281 (8): 4787-4794. 10.1074/джбк.М508657200
КАС пабмед Статья Google Scholar
Кишида М., Кояма С., Кисида С., Мацубара К., Накашима С., Хигано К., Такада Р., Такада С., Кикучи А.: Аксин предотвращает индуцированное Wnt-3a накопление β-катенина. Онкоген. 1999, 18 (4): 979-985. 10.1038/sj.onc.1202388
CAS пабмед Статья Google Scholar
Даджани Р., Фрейзер Э., Роу С.М., Йео М., Гуд В.М., Томпсон В., Дейл Т.С., Перл Л.Х.: Структурная основа для рекрутирования киназы гликогенсинтазы 3β в каркасный комплекс аксин-АРС.EMBO J. 2003, 22 (3): 494-501. 10.1093/emboj/cdg068
Центральный PubMed КАС пабмед Статья Google Scholar
Willert K, Shibamoto S, Nusse R: Wnt-индуцированное дефосфорилирование Axin высвобождает β-катенин из комплекса Axin. Гены Дев. 1999, 13 (14): 1768-1773. 10.1101/гад.14.13.1768
Гены Дев. 1999, 13 (14): 1768-1773. 10.1101/гад.14.13.1768
PubMed Central КАС пабмед Статья Google Scholar
Луо В., Петерсон А., Гарсия Б.А., Кумбс Г., Кофаль Б., Генрих Р., Шабановиц Дж., Хант Д.Ф., Йост Х.Дж., Виршуп Д.М.: Протеинфосфатаза 1 регулирует сборку и функцию комплекса деградации β-катенина. EMBO J. 2007, 26 (6): 1511-1521. 10.1038/sj.emboj.7601607
PubMed Central КАС пабмед Статья Google Scholar
Strovel ET, Wu D, Sussman DJ: Протеинфосфатаза 2Cα дефосфорилирует аксин и активирует LEF-1-зависимую транскрипцию.Дж. Биол. Хим. 2000, 275 (4): 2399-2403. 10.1074/джбк.275.4.2399
КАС пабмед Статья Google Scholar
Hinoi T, Yamamoto H, Kishida M, Takada S, Kishida S, Kikuchi A: Комплексное образование продукта гена аденоматозного полипоза кишечной палочки и Axin облегчает гликогенсинтазкиназу-3β-зависимое фосфорилирование β-катенина и подавление регулирует β-катенин. Дж. Биол. Хим. 2000, 275 (44): 34399-34406.
Дж. Биол. Хим. 2000, 275 (44): 34399-34406.
КАС пабмед Статья Google Scholar
Huang H, He X: Передача сигналов Wnt/β-катенина: новые (и старые) игроки и новые идеи. Curr Opin Cell Biol. 2008, 20 (2): 119-125. 10.1016/j.ceb.2008.01.009
PubMed Central КАС пабмед Статья Google Scholar
Angers S, Moon RT: Проксимальные события в передаче сигнала Wnt. Nat Rev Mol Cell Biol. 2009, 10 (7): 468-477.
КАС пабмед Статья Google Scholar
Tamai K, Zeng X, Liu C, Zhang X, Harada Y, Chang Z, He X: Механизм активации корецептора Wnt. Мол Ячейка. 2004, 13 (1): 149-156. 10.1016/С1097-2765(03)00484-2
КАС пабмед Статья Google Scholar
Niehrs C, Shen J: Регуляция фосфорилирования Lrp6. Cell Mol Life Sci. 2010, 67 (15): 2551-2562. 10.1007/s00018-010-0329-3
Cell Mol Life Sci. 2010, 67 (15): 2551-2562. 10.1007/s00018-010-0329-3
КАС пабмед Статья Google Scholar
Pan W, Choi SC, Wang H, Qin Y, Volpicelli-Daley L, Swan L, Lucast L, Khoo C, Zhang X, Li L, Abrams CS, Sokol SY, Wu D: Wnt3a-опосредованное образование фосфатидилинозитола 4, 5-бисфосфат регулирует фосфорилирование LRP6. Наука. 2008, 321 (5894): 1350-1353. 10.1126/science.1160741
PubMed Central КАС пабмед Статья Google Scholar
Цзэн Х, Хуан Х, Тамай К, Чжан Х, Харада Й, Йокота С, Алмейда К, Ван Дж, Доубл Б, Вуджетт Дж, Виншоу-Борис А, Се Дж. К., Хе Х: Инициация Wnt передача сигналов: контроль фосфорилирования/активации корецептора Wnt Lrp6 посредством функций взъерошенных, растрепанных и аксинов.Разработка. 2008, 135 (2): 367-375.
КАС пабмед Статья Google Scholar
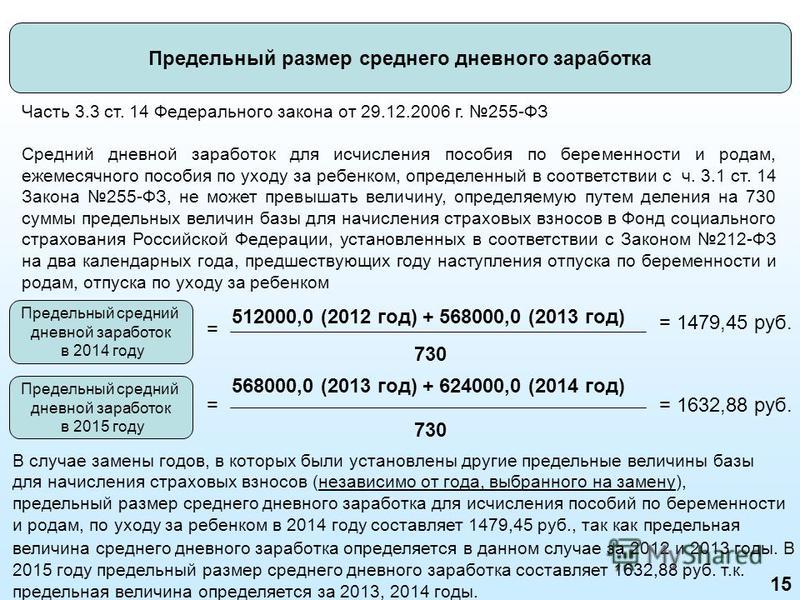
Smalley MJ, Sara E, Paterson H, Naylor S, Cook D, Jayatilake H, Fryer LG, Hutchinson L, Fry MJ, Dale TC: Взаимодействие белков Axin и Dvl-2 регулирует Dvl-2-стимулированные TCF-зависимая транскрипция. EMBO J. 1999, 18 (10): 2823-2835. 10.1093/emboj/18.10.2823
Центральный пабмед КАС пабмед Статья Google Scholar
Kishida S, Yamamoto H, Hino S, Ikeda S, Kishida M, Kikuchi A: домены DIX Dvl и аксина необходимы для белковых взаимодействий и их способности регулировать стабильность β-катенина. Мол Селл Биол. 1999, 19 (6): 4414-4422.
Центральный пабмед КАС пабмед Статья Google Scholar
Fiedler M, Mendoza-Topaz C, Rutherford TJ, Mieszcchanek J, Bienz M: Disheveled взаимодействует с интерфейсом полимеризации домена DIX Axin, чтобы препятствовать его функции по подавлению β-катенина.Proc Natl Acad Sci USA. 2011, 108 (5): 1937-1942. 10.1073/pnas.1017063108
10.1073/pnas.1017063108
PubMed Central КАС пабмед Статья Google Scholar
Mao J, Wang J, Liu B, Pan W, Farr Iii GH, Flynn C, Yuan H, Takada S, Kimelman D, Li L, Wu D: Белок-5, связанный с рецептором липопротеинов низкой плотности. связывается с Axin и регулирует канонический сигнальный путь Wnt. Мол Ячейка. 2001, 7 (4): 801-809. 10.1016/С1097-2765(01)00224-6
КАС пабмед Статья Google Scholar
Bilić J, Huang YL, Davidson G, Zimmermann T, Cruciat CM, Bienz M, Niehrs C: Wnt индуцирует сигналосомы LRP6 и способствует фосфорилированию LRP6, зависимому от растрепанности. Наука. 2007, 316 (5831): 1619-1622. 10.1126/наука.1137065
PubMed Статья КАС Google Scholar
Schwarz-Romond T., Fiedler M., Shibata N., Butler PJG, Kikuchi A., Higuchi Y., Bienz M. Домен DIX Disheveled обеспечивает передачу сигналов Wnt посредством динамической полимеризации.Nat Struct Mol Biol. 2007, 14 (6): 484-492. 10.1038/нсмб1247
Домен DIX Disheveled обеспечивает передачу сигналов Wnt посредством динамической полимеризации.Nat Struct Mol Biol. 2007, 14 (6): 484-492. 10.1038/нсмб1247
КАС пабмед Статья Google Scholar
Schwarz-Romond T, Metcalfe C, Bienz M: Динамическое рекрутирование аксина с помощью белковых сборок Disheveled. Дж. Клеточные науки. 2007, 120 (14): 2402-2412. 10.1242/jcs.002956
CAS пабмед Статья Google Scholar
Lee JS, Ishimoto A, Yanagawa S: Характеристика растрепанных белков мыши (Dvl) в сигнальном пути Wnt/Wingless.Дж. Биол. Хим. 1999, 274 (30): 21464-21470. 10.1074/джбк.274.30.21464
КАС пабмед Статья Google Scholar
González-Sancho JM, Brennan KR, Castelo-Soccio LA, Brown AM: Белки Wnt индуцируют беспорядочное фосфорилирование посредством LRP5/6-независимого механизма, независимо от их способности стабилизировать β-катенин. Мол Селл Биол. 2004, 24 (11): 4757-4768. 10.1128/MCB.24.11.4757-4768.2004
Мол Селл Биол. 2004, 24 (11): 4757-4768. 10.1128/MCB.24.11.4757-4768.2004
PubMed Central пабмед Статья КАС Google Scholar
Doble BW, Patel S, Wood GA, Kockeritz LK, Woodgett JR: Функциональная избыточность GSK-3α и GSK-3β в передаче сигналов Wnt/β-катенина, показанная с использованием аллельной серии линий эмбриональных стволовых клеток. Ячейка Дев. 2007, 12 (6): 957-971. 10.1016/j.devcel.2007.04.001
Центральный PubMed КАС пабмед Статья Google Scholar
Коэн П., Фрейм S: Возрождение GSK3. Nat Rev Mol Cell Biol. 2001, 2 (10): 769-776. 10.1038/35096075
КАС пабмед Статья Google Scholar
Frame S, Cohen P, Biondi RM: Общий сайт связывания фосфатов объясняет уникальную субстратную специфичность GSK3 и его инактивацию путем фосфорилирования. Мол Ячейка. 2001, 7 (6): 1321-1327. 10.1016/С1097-2765(01)00253-2
2001, 7 (6): 1321-1327. 10.1016/С1097-2765(01)00253-2
КАС пабмед Статья Google Scholar
Даджани Р., Фрейзер Э., Роу С.М., Янг Н., Гуд В., Дейл Т.С., Перл Л.Х.: Кристаллическая структура киназы гликогенсинтазы 3β: структурная основа субстратной специфичности и аутоингибирования, примированных фосфатом.Клетка. 2001, 105 (6): 721-732. 10.1016/S0092-8674(01)00374-9
КАС пабмед Статья Google Scholar
Fukumoto S, Hsieh CM, Maemura K, Layne MD, Yet SF, Lee KH, Matsui T, Rosenzweig A, Taylor WG, Rubin JS, Perrella MA, Lee ME: участие Akt в сигнальном пути Wnt через Растрепанный. Дж. Биол. Хим. 2001, 276 (20): 17479-17483. 10.1074/джбк.К000880200
КАС пабмед Статья Google Scholar
Desbois-Mouthon C, Cadoret A, Blivet-Van Eggelpoël MJ, Bertrand F, Cherqui G, Perret C, Capeau J: Инсулин и IGF-1 стимулируют путь β-катенина посредством двух сигнальных каскадов, включающих ингибирование GSK-3β и активацию Ras . Онкоген. 2001, 20 (2): 252-259. 10.1038/sj.onc.1204064
Онкоген. 2001, 20 (2): 252-259. 10.1038/sj.onc.1204064
CAS пабмед Статья Google Scholar
Ding VW, Chen RH, McCormick F: Дифференциальная регуляция киназы 3β гликогенсинтазы с помощью инсулина и передачи сигналов Wnt.Дж. Биол. Хим. 2000, 275 (42): 32475-32481.
КАС пабмед Статья Google Scholar
McManus EJ, Sakamoto K, Armit LJ, Ronaldson L, Shpiro N, Marquez R, Alessi DR: Роль, которую играет фосфорилирование GSK3 в передаче сигналов инсулина и Wnt, определяется анализом нокина. EMBO J. 2005, 24 (8): 1571-1583. 10.1038/sj.emboj.7600633
PubMed Central КАС пабмед Статья Google Scholar
Bikkavilli RK, Feigin ME, Malbon CC: митоген-активируемая протеинкиназа p38 регулирует каноническую передачу сигналов Wnt-β-катенин путем инактивации GSK3β. Дж. Клеточные науки. 2008, 121 (21): 3598-3607. 10.1242/JCS.032854
Клеточные науки. 2008, 121 (21): 3598-3607. 10.1242/JCS.032854
CAS пабмед Статья Google Scholar
Wu ZQ, Brabletz T, Fearon E, Willis AL, Hu CY, Li XY, Weiss SJ: Канонический супрессор Wnt, Axin2, способствует онкогенной активности карциномы толстой кишки. Proc Natl Acad Sci U S A.2012, 109 (28): 11312-11317. 10.1073/pnas.1203015109
PubMed Central КАС пабмед Статья Google Scholar
Itoh K, Tang TL, Neel BG, Sokol SY: Специфическая модуляция судеб эктодермальных клеток у эмбрионов Xenopus киназой гликогенсинтазы. Разработка. 1995, 121 (12): 3979-3988.
КАС пабмед Google Scholar
Buescher JL, Phiel CJ: Некаталитический домен киназы гликогенсинтазы-3 (GSK-3) необходим для активности.Дж. Биол. Хим. 2010, 285 (11): 7957-7963. 10.1074/jbc.M109.0
Центральный пабмед КАС пабмед Статья Google Scholar
Fraser E, Young N, Dajani R, Franca-Koh J, Ryves J, Williams RSB, Yeo M, Webster MT, Richardson C, Smalley MJ, Pearl LH, Harwood A, Dale TC: Идентификация Область связывания аксина и Frat киназы-3 гликогенсинтазы. Дж. Биол. Хим. 2002, 277 (3): 2176-2185. 10.1074/джбк.М109462200
КАС пабмед Статья Google Scholar
Zhang Y, Qiu WJ, Liu DX, Neo SY, He X, Lin SC: Дифференциальные молекулярные сборки лежат в основе двойной функции Axin в модулировании путей WNT и JNK. Дж. Биол. Хим. 2001, 276 (34): 32152-32159. 10.1074/джбк.М104451200
КАС пабмед Статья Google Scholar
Roose J, Molenaar M, Peterson J, Hurenkamp J, Brantjes H, Moerer P, van de Wetering M, Destree O, Clevers H: эффектор Xenopus Wnt XTcf-3 взаимодействует с репрессорами транскрипции, связанными с Groucho.Природа. 1998, 395 (6702): 608-612. 10.1038/26989
КАС пабмед Статья Google Scholar
Arce L, Pate K, Waterman M: Граучо связывает две консервативные области LEF-1 для HDAC-зависимой репрессии. БМК Рак. 2009, 9 (1): 159. 10.1186/1471-2407-9-159
PubMed Central пабмед Статья КАС Google Scholar
Daniels DL, Weis WI: β-катенин напрямую вытесняет репрессоры Groucho/TLE из Tcf/Lef в Wnt-опосредованной активации транскрипции.Nat Struct Mol Biol. 2005, 12 (4): 364-371. 10.1038/нсмб912
КАС пабмед Статья Google Scholar
Hsu SC, Galceran J, Grosschedl R: Модуляция регуляции транскрипции с помощью LEF-1 в ответ на передачу сигналов Wnt-1 и связь с β-катенином. Мол Селл Биол. 1998, 18 (8): 4807-4818.
Центральный пабмед КАС пабмед Статья Google Scholar
Гершко А., Цехановер А.: Система убиквитина. Анну Рев Биохим. 1998, 67 (1): 425-479. 10.1146/annurev.biochem.67.1.425
10.1146/annurev.biochem.67.1.425
CAS пабмед Статья Google Scholar
Пикарт С.М., Эддинс М.Дж.: Убиквитин: структуры, функции, механизмы. Биохим Биофиз Акта. 2004, 1695 (1–3): 55–72.
КАС пабмед Статья Google Scholar
Glickman MH, Ciechanover A: Убиквитин-протеасомный протеолитический путь: разрушение ради строительства. Physiol Rev. 2002, 82 (2): 373-428.
КАС пабмед Статья Google Scholar
Mukhopadhyay D, Riezman H: Независимые от протеасом функции убиквитина в эндоцитозе и передаче сигналов. Наука. 2007, 315 (5809): 201-205. 10.1126/наука.1127085
CAS пабмед Статья Google Scholar
Schnell JD, Hicke L: Нетрадиционные функции убиквитина и убиквитин-связывающих белков. Дж. Биол. Хим. 2003, 278 (38): 35857-35860. 10.1074/джбк.Р300018200
Дж. Биол. Хим. 2003, 278 (38): 35857-35860. 10.1074/джбк.Р300018200
КАС пабмед Статья Google Scholar
Chen ZJ, Sun LJ: Непротеолитические функции убиквитина в клеточной передаче сигналов. Мол Ячейка. 2009, 33 (3): 275-286. 10.1016/j.molcel.2009.01.014
CAS пабмед Статья Google Scholar
Komander D: Возникающая сложность убиквитинирования белков. Биохим Сок Транс. 2009, 37 (часть 5): 937-953.
КАС пабмед Статья Google Scholar
Kulathu Y, Komander D: Атипичное убиквитилирование — неизведанный мир полиубиквитина за пределами связей Lys48 и Lys63. Nat Rev Mol Cell Biol. 2012, 13 (8): 508-523. 10.1038/nrm3394
КАС пабмед Статья Google Scholar
Икеда Ф., Дикич И.: Атипичные цепи убиквитина: новые молекулярные сигналы. EMBO Rep. 2008, 9 (6): 536-542. 10.1038/embor.2008.93
EMBO Rep. 2008, 9 (6): 536-542. 10.1038/embor.2008.93
PubMed Central КАС пабмед Статья Google Scholar
Кирисако Т., Камей К., Мурата С., Като М., Фукумото Х., Кани М., Сано С., Токунага Ф., Танака К., Иваи К.: Комплекс убиквитинлигазы собирает линейные полиубиквитиновые цепи. EMBO J. 2006, 25 (20): 4877-4887. 10.1038/sj.emboj.7601360
PubMed Central КАС пабмед Статья Google Scholar
Токунага Ф., Саката Си, Саэки Ю., Сатоми Ю., Кирисако Т., Камей К., Накагава Т., Като М., Мурата С., Ямаока С., Ямамото М., Акира С., Такао Т., Танака К., Иваи К.: Участие линейного полиубиквитилирования NEMO в активации NF-κB. Nat Cell Biol. 2009, 11 (2): 123-132. 10.1038/ncb1821
КАС пабмед Статья Google Scholar
Соркин А. Регуляция эндоцитарного транспорта рецепторов и транспортеров путем убиквитинирования: возможная роль в нейродегенеративных заболеваниях. Внутриклеточный трафик и нейродегенеративные расстройства. Под редакцией: Джордж-Хислоп П.С., Мобли В.К., Кристен Ю. 2009, 141-155. Берлин, Гейдельберг: Springer.
Внутриклеточный трафик и нейродегенеративные расстройства. Под редакцией: Джордж-Хислоп П.С., Мобли В.К., Кристен Ю. 2009, 141-155. Берлин, Гейдельберг: Springer.
Глава Google Scholar
Кравцова-Иванцив Ю., Чехановер А.: Неканонические сигналы на основе убиквитина для протеасомной деградации. Дж. Клеточные науки. 2012, 125 (3): 539-548. 10.1242/jcs.0
CAS пабмед Статья Google Scholar
Фукс С.Ю., Шпигельман В.С., Суреш Кумар К.Г.: Многоликость убиквитинлигазы β-TrCP E3: отражение в волшебном зеркале рака. Онкоген. 2004, 23 (11): 2028-2036. 10.1038/sj.onc.1207389
CAS пабмед Статья Google Scholar
Панченко М.В., Чжоу М.И., Коэн Х.Т.: партнер фон Хиппель-Линдау Jade-1 Является коактиватором транскрипции, связанным с активностью гистон-ацетилтрансферазы. Дж. Биол. Хим. 2004, 279 (53): 56032-56041.10.1074/джбк.М410487200
Биол. Хим. 2004, 279 (53): 56032-56041.10.1074/джбк.М410487200
КАС пабмед Статья Google Scholar
Zhou MI, Wang H, Ross JJ, Kuzmin I, Xu C, Cohen HT: Супрессор опухоли фон Хиппеля-Линдау стабилизирует новый растительный гомеодоменовый белок Jade-1. Дж. Биол. Хим. 2002, 277 (42): 39887-39898. 10.1074/джбк.М205040200
КАС пабмед Статья Google Scholar
Behrens J, Jerchow BA, Würtele M, Grimm J, Asbrand C, Wirtz R, Kühl M, Wedlich D, Birchmeier W: Функциональное взаимодействие гомолога аксина, кондуктина, с β-катенином, APC и ГСК3β.Наука. 1998, 280 (5363): 596-599. 10.1126/наука.280.5363.596
CAS пабмед Статья Google Scholar
Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P: Понижающая регуляция β-катенина аксином человека и его связь с супрессором опухоли APC, β-катенином и GSK3β. Карр Биол. 1998, 8 (10): 573-581. 10.1016/С0960-9822(98)70226-С
Карр Биол. 1998, 8 (10): 573-581. 10.1016/С0960-9822(98)70226-С
КАС пабмед Статья Google Scholar
Lee E, Salic A, Krüger R, Heinrich R, Kirschner MW: роли APC и Axin, полученные на основе экспериментального и теоретического анализа пути Wnt. PLoS биол. 2003, 1 (1): e10. 10.1371/journal.pbio.0000010
PubMed Central пабмед Статья Google Scholar
Salic A, Lee E, Mayer L, Kirschner MW: Контроль стабильности β-катенина: восстановление цитоплазматических стадий пути Wnt в экстрактах яиц Xenopus.Мол Ячейка. 2000, 5 (3): 523-532. 10.1016/С1097-2765(00)80446-3
КАС пабмед Статья Google Scholar
Lui TT, Lacroix C, Ahmed SM, Goldenberg SJ, Leach CA, Daulat AM, Angers S: Убиквитин-специфическая протеаза USP34 регулирует стабильность аксина и передачу сигналов Wnt/β-катенина. Мол Селл Биол. 2011, 31 (10): 2053-2065. 10.1128/MCB.01094-10
Мол Селл Биол. 2011, 31 (10): 2053-2065. 10.1128/MCB.01094-10
Центральный пабмед КАС пабмед Статья Google Scholar
Petroski MD, Deshaies RJ: Функция и регуляция убиквитинлигазы cullin-RING. Nat Rev Mol Cell Biol. 2005, 6 (1): 9-20. 10.1038/nrm1547
КАС пабмед Статья Google Scholar
Papkoff J, Rubinfeld B, Schryver B, Polakis P: Wnt-1 регулирует свободные пулы катенинов и стабилизирует комплексы APC-катенин. Мол Селл Биол. 1996, 16 (5): 2128-2134.
Центральный пабмед КАС пабмед Статья Google Scholar
Gao C, Chen YG: Растрепанный: узел сигнализации Wnt. Сотовый сигнал. 2010, 22 (5): 717-727. 10.1016/j.cellsig.2009.11.021
CAS пабмед Статья Google Scholar
Habas R: сигнализация Canonical Wnt: неожиданный новый игрок. Ячейка Дев. 2006, 11 (2): 138-139. 10.1016/j.devcel.2006.07.009
Ячейка Дев. 2006, 11 (2): 138-139. 10.1016/j.devcel.2006.07.009
CAS пабмед Статья Google Scholar
Тауриелло Д.В., Морис М.М.: Различные роли убиквитина в регуляции пути Wnt. Клеточный цикл. 2010, 9 (18): 3724-3733.
Артикул КАС Google Scholar
González-Sancho JM, Greer YE, Abrahams CL, Takigawa Y, Baljinnyam B, Lee KH, Lee KS, Rubin JS, Brown AM: Функциональные последствия Wnt-индуцированного фосфорилирования взлохмаченного 2 в канонической и неканонической передаче сигналов Wnt . Дж. Биол. Хим. 2013, 288 (13): 9428-9437.10.1074/jbc.M112.448480
Центральный пабмед пабмед Статья КАС Google Scholar
Cadigan KM, Fish MP, Rulifson EJ, Nusse R: Подавление Wingless экспрессии Drosophila frizzled 2 формирует градиент морфогена Wingless в крыле. Клетка. 1998, 93 (5): 767-777. 10.1016/S0092-8674(00)81438-5
1998, 93 (5): 767-777. 10.1016/S0092-8674(00)81438-5
КАС пабмед Статья Google Scholar
Zhang J, Li Y, Liu Q, Lu W, Bu G: активация передачи сигналов Wnt и гиперплазия молочной железы у трансгенных мышей MMTV-LRP6: значение для онкогенеза рака молочной железы. Онкоген. 2009, 29 (4): 539-549.
Центральный пабмед КАС пабмед Статья Google Scholar
Haglund K, Dikic I: Роль убиквитилирования в эндоцитозе рецепторов и эндосомальной сортировке. Дж. Клеточные науки. 2012, 125 (2): 265-275. 10.1242/JCS.0
CAS пабмед Статья Google Scholar
Tran H, Polakis P: Обратимая модификация аденоматозного полипоза толстой кишки (APC) полиубиквитином, связанным с K63, регулирует сборку и активность комплекса разрушения β-катенина. Дж. Биол. Хим. 2012, 287 (34): 28552-28563. 10.1074/jbc.M112.387878
10.1074/jbc.M112.387878
Центральный пабмед КАС пабмед Статья Google Scholar
Бигнелл Г.Р., Уоррен В., Сил С., Такахаши М., Рэпли Э., Барфут Р., Грин Х., Браун С., Биггс П.Дж., Лахани С.Р., Джонс С., Хансен Дж., Блэр Э., Хофманн Б., Зиберт Р. , Тернер Г., Эванс Д.Г., Шрандер-Штумпель С., Бимер Ф.А., ван Ден Оувеланд А., Галлей Д., Делпеш Б., Кливленд М.Г., Ли И., Лейсти Дж., Расмуссен С.: Идентификация гена-супрессора опухоли семейного цилиндроматоза.Нат Жене. 2000, 25 (2): 160-165. 10.1038/76006
КАС пабмед Статья Google Scholar
Jürgen Dohmen R: Модификация белка SUMO. Биохим Биофиз Акта. 2004, 1695 (1–3): 113–131.
ПабМед Статья КАС Google Scholar
Gareau JR, Lima CD: Путь SUMO: новые механизмы, формирующие специфичность, конъюгацию и распознавание.Nat Rev Mol Cell Biol. 2010, 11 (12): 861-871. 10.1038/nrm3011
Центр PubMed КАС пабмед Статья Google Scholar
Yeh ETH: СУМОилирование и Де-СУМОилирование: борьба с жизненными процессами. Дж. Биол. Хим. 2009, 284 (13): 8223-8227. 10.1074/jbc.R800050200
Центральный пабмед КАС пабмед Статья Google Scholar
Matunis MJ, Coutavas E, Blobel G: Новая убиквитин-подобная модификация модулирует разделение Ran-GTPase-активирующего белка RanGAP1 между цитозолем и комплексом ядерной поры.Джей Селл Биол. 1996, 135 (6): 1457-1470. 10.1083/джкб.135.6.1457
КАС пабмед Статья Google Scholar
Кершер О: Перекресток СУМО — какая у тебя функция? Новое понимание через взаимодействующие с СУМО мотивы. EMBO Rep. 2007, 8 (6): 550-555. 10.1038/sj.embor.7400980
PubMed Central КАС пабмед Статья Google Scholar
Johnson ES: Модификация белка с помощью SUMO.Анну Рев Биохим. 2004, 73 (1): 355-382. 10.1146/annurev.biochem.73.011303.074118
CAS пабмед Статья Google Scholar
Kadoya T, Kishida S, Fukui A, Hinoi T, Michiue T, Asashima M, Kikuchi A: ингибирование сигнального пути Wnt новым аксин-связывающим белком. Дж. Биол. Хим. 2000, 275 (47): 37030-37037. 10.1074/джбк.М005984200
КАС пабмед Статья Google Scholar
Кадоя Т., Ямамото Х., Сузуки Т., Юкита А., Фукуи А., Мичиуэ Т., Асахара Т., Танака К., Асашима М., Кикучи А.: Десумоилирующая активность Аксама, нового аксин-связывающего белка, участвует в подавлении β- катенин. Мол Селл Биол. 2002, 22 (11): 3803-3819. 10.1128/MCB.22.11.3803-3819.2002
PubMed Central КАС пабмед Статья Google Scholar
Kim MJ, Chia IV, Costantini F: Сайты-мишени SUMOylation на C-конце защищают Axin от убиквитинирования и обеспечивают стабильность белка.FASEB J. 2008, 22 (11): 3785-3794. 10.1096/fj.08-113910
Центральный пабмед КАС пабмед Статья Google Scholar
Li J, Wang CY: TBL1-TBLR1 и β-катенин рекрутируют друг друга на промотор гена-мишени Wnt для активации транскрипции и онкогенеза. Nat Cell Biol. 2008, 10 (2): 160-169. 10.1038/нкб1684
КАС пабмед Статья Google Scholar
Choi HK, Choi KC, Yoo JY, Song M, Ko Suk J, Kim Chul H, Ahn JH, Chun KH, Yook Jong I, Yoon HG: Обратимое СУМОилирование TBL1-TBLR1 регулирует опосредованную β-катенином передачу сигналов Wnt. Мол Ячейка. 2011, 43 (2): 203-216. 10.1016/j.molcel.2011.05.027
CAS пабмед Статья Google Scholar
Li M, Brooks CL, Wu-Baer F, Chen D, Baer R, Gu W: моно- и полиубиквитинирование: дифференциальный контроль судьбы p53 с помощью Mdm2.Наука. 2003, 302 (5652): 1972-1975. 10.1126/наука.10
CAS пабмед Статья Google Scholar
Юрченко В., Сюэ З., Садофски М.Дж.: Модификация SUMO человеческого XRCC4 регулирует его локализацию и функцию в репарации двухцепочечных разрывов ДНК. Мол Селл Биол. 2006, 26 (5): 1786-1794. 10.1128/MCB.26.5.1786-1794.2006
PubMed Central КАС пабмед Статья Google Scholar
Huang J, Yan J, Zhang J, Zhu S, Wang Y, Shi T, Zhu C, Chen C, Liu X, Cheng J, Mustelin T, Feng GS, Chen G, Yu J: модификация PTEN SUMO1 регулирует онкогенез путем контролируя его связь с плазматической мембраной. Нац коммун. 2012, 3: 911.
ПабМед Статья КАС Google Scholar
Kubota Y, O’Grady P, Saito H, Takekawa M: Онкогенный Ras отменяет SUMOylation MEK, который подавляет путь ERK и трансформацию клеток.Nat Cell Biol. 2011, 13 (3): 282-291. 10.1038/ncb2169
КАС пабмед Статья Google Scholar
Carter S, Vousden KH: слияния p53-Ubl как модели убиквитинирования, сумоилирования и недилирования p53. Клеточный цикл. 2008, 7 (16): 2519-2528. 10.4161/cc.7.16.6422
CAS пабмед Статья Google Scholar
Лундби А., Лаге К., Вейнерт Брайан Т., Беккер-Дженсен Дорте Б., Сечер А., Сковгаард Т., Келструп Кристиан Д., Дмитриев А., Чоудхари С., Лундби С., Олсен Дж. В.: Протеомный анализ сайтов ацетилирования лизина в тканях крыс выявляет органную специфичность и субклеточные паттерны.Cell Rep. 2012, 2 (2): 419-431. 10.1016/j.celrep.2012.07.006
PubMed Central КАС пабмед Статья Google Scholar
Yang XJ: Разнообразное суперсемейство лизин-ацетилтрансфераз и их роль при лейкемии и других заболеваниях. Нуклеиновые Кислоты Res. 2004, 32 (3): 959-976. 10.1093/nar/gkh352
Центральный пабмед КАС пабмед Статья Google Scholar
Guan KL, Xiong Y: Регуляция промежуточного метаболизма путем ацетилирования белков. Тенденции биохимических наук. 2011, 36 (2): 108-116. 10.1016/j.tibs.2010.09.003
PubMed Central КАС пабмед Статья Google Scholar
Sterner DE, Berger SL: Ацетилирование гистонов и факторов, связанных с транскрипцией. Microbiol Mol Biol Rev. 2000, 64 (2): 435-459. 10.1128/MMBR.64.2.435-459.2000
PubMed Central КАС пабмед Статья Google Scholar
Струль К.: Ацетилирование гистонов и механизмы регуляции транскрипции. Гены Дев. 1998, 12 (5): 599-606. 10.1101/гад.12.5.599
КАС пабмед Статья Google Scholar
Strahl BD, Allis CD: Язык ковалентных модификаций гистонов. Природа. 2000, 403 (6765): 41-45. 10.1038/47412
КАС пабмед Статья Google Scholar
Spange S, Wagner T, Heinzel T, Krämer OH: Ацетилирование негистоновых белков модулирует клеточную передачу сигналов на нескольких уровнях. Int J Biochem Cell Biol. 2009, 41 (1): 185-198. 10.1016/j.biocel.2008.08.027
CAS пабмед Статья Google Scholar
Глозак М.А., Сенгупта Н., Чжан X, Сето Э.: Ацетилирование и деацетилирование негистоновых белков. Ген. 2005, 363: 15-23.
КАС пабмед Статья Google Scholar
Чоудхари С., Кумар С., Гнад Ф., Нильсен М.Л., Рехман М., Вальтер Т.С., Олсен Дж.В., Манн М.: Ацетилирование лизина нацелено на белковые комплексы и совместно регулирует основные клеточные функции. Наука. 2009, 325 (5942): 834-840. 10.1126/наука.1175371
CAS пабмед Статья Google Scholar
Firestein R, Blander G, Michan S, Oberdoerffer P, Ogino S, Campbell J, Bhimavarapu A, Luikenhuis S, de Cabo R, Fuchs C, Hahn WC, Guarente LP, Sinclair DA: Деацетилаза SIRT1 подавляет туморогенез кишечника и рост рака толстой кишки.ПЛОС Один. 2008, 3 (4): e2020. 10.1371/journal.pone.0002020
PubMed Central пабмед Статья КАС Google Scholar
Ким М.Ю., Чжан Т., Краус В.Л.: Поли(АДФ-рибозилирование) с помощью PARP-1: «PAR-укладка» NAD+ в ядерный сигнал. Гены Дев. 2005, 19 (17): 1951-1967. 10.1101/гад.1331805
КАС пабмед Статья Google Scholar
Hassa PO, Haenni SS, Elser M, Hottiger MO: Реакции ядерного АДФ-рибозилирования в клетках млекопитающих: где мы сегодня и куда мы идем?.Microbiol Mol Biol Rev. 2006, 70 (3): 789-829. 10.1128/MMBR.00040-05
PubMed Central КАС пабмед Статья Google Scholar
Schreiber V, Dantzer F, Ame JC, de Murcia G: Поли(АДФ-рибоза): новые функции старой молекулы. Nat Rev Mol Cell Biol. 2006, 7 (7): 517-528. 10.1038/nrm1963
CAS пабмед Статья Google Scholar
James RG, Davidson KC, Bosch KA, Biechele TL, Robin NC, Taylor RJ, Major MB, Camp ND, Fowler K, Martins TJ, Moon RT: WIKI4, новый ингибитор танкиразы и передачи сигналов Wnt/β-катенина. ПЛОС Один. 2012, 7 (12): e50457. 10.1371/journal.pone.0050457
PubMed Central КАС пабмед Статья Google Scholar
Хантер Т. Эпоха перекрестных помех: фосфорилирование, убиквитинирование и прочее. Мол Ячейка. 2007, 28 (5): 730-738.10.1016/j.molcel.2007.11.019
CAS пабмед Статья Google Scholar
Yang XJ, Seto E: Ацетилирование лизина: систематизированные перекрестные помехи с другими посттрансляционными модификациями. Мол Ячейка. 2008, 31 (4): 449-461. 10.1016/j.molcel.2008.07.002
PubMed Central КАС пабмед Статья Google Scholar
Guo Z, Kanjanapangka J, Liu N, Liu S, Liu C, Wu Z, Wang Y, Loh T, Kowolik C, Jamsen J, Zhou M, Truong K, Chen Y, Zheng L, Shen B : Последовательные посттрансляционные модификации программируют деградацию FEN1 во время прогрессирования клеточного цикла.Мол Ячейка. 2012, 47 (3): 444-456. 10.1016/j.molcel.2012.05.042
PubMed Central КАС пабмед Статья Google Scholar
Мурр Р.: Взаимодействие между различными эпигенетическими модификациями и механизмами. Достижения в области генетики, том 70. Под редакцией: Зденко Х., Тошиказу У. 2010, 101–141. Уолтем, Массачусетс: Academic Press.
Google Scholar
Wang Z, Michaud GA, Cheng Z, Zhang Y, Hinds TR, Fan E, Cong F, Xu W: Распознавание фрагмента изо-АДФ-рибозы в поли(АДФ-рибозе) доменами WWE предполагает общий механизм поли (АДФ-рибозил)ирование-зависимое убиквитинирование. Гены Дев. 2012, 26 (3): 235-240. 10.1101/gad.182618.111
PubMed Central пабмед Статья КАС Google Scholar
Kang HC, Lee YI, Shin JH, Andrabi SA, Chi Z, Gagné JP, Lee Y, Ko HS, Lee BD, Poirier GG, Dawson VL, Dawson TM: Iduna представляет собой поли(АДФ-рибозу). ) (PAR)-зависимая убиквитинлигаза E3, которая регулирует повреждение ДНК.Proc Natl Acad Sci USA. 2011, 108 (34): 14103-14108. 10.1073/pnas.1108799108
PubMed Central КАС пабмед Статья Google Scholar
Nusse R, Varmus H: Три десятилетия Wnts: личный взгляд на развитие научной области. EMBO J. 2012, 31 (12): 2670-2684. 10.1038/emboj.2012.146
PubMed Central КАС пабмед Статья Google Scholar
Anastas JN, Moon RT: сигнальные пути WNT как терапевтические мишени при раке. Нат Рев Рак. 2013, 13 (1): 11-26.
КАС пабмед Статья Google Scholar
Polakis P: Лекарственное воздействие на передачу сигналов Wnt при раке. EMBO J. 2012, 31 (12): 2737-2746. 10.1038/emboj.2012.126
PubMed Central КАС пабмед Статья Google Scholar
%PDF-1.4 % 925 0 объект > эндообъект внешняя ссылка 925 124 0000000016 00000 н 0000005659 00000 н 0000005867 00000 н 0000005903 00000 н 0000006429 00000 н 0000006583 00000 н 0000006740 00000 н 0000006894 00000 н 0000007051 00000 н 0000007205 00000 н 0000007362 00000 н 0000007519 00000 н 0000007676 00000 н 0000007833 00000 н 0000007988 00000 н 0000008637 00000 н 0000010768 00000 н 0000011152 00000 н 0000011704 00000 н 0000012127 00000 н 0000012999 00000 н 0000013864 00000 н 0000014787 00000 н 0000015658 00000 н 0000016852 00000 н 0000018045 00000 н 0000018527 00000 н 0000018587 00000 н 0000022715 00000 н 0000023159 00000 н 0000023440 00000 н 0000023882 00000 н 0000024383 00000 н 0000024948 00000 н 0000025884 00000 н 0000026752 00000 н 0000027222 00000 н 0000027922 00000 н 0000029106 00000 н 0000029190 00000 н 0000029668 00000 н 0000030236 00000 н 0000030890 00000 н 0000031358 00000 н 0000032555 00000 н 0000033216 00000 н 0000035935 00000 н 0000036754 00000 н 0000037521 00000 н 0000038143 00000 н 0000038965 00000 н 0000039758 00000 н 0000047427 00000 н 0000055696 00000 н 0000062314 00000 н 0000068488 00000 н 0000074498 00000 н 0000081482 00000 н 0000081554 00000 н 0000081770 00000 н 0000081870 00000 н 0000081993 00000 н 0000082161 00000 н 0000082325 00000 н 0000082478 00000 н 0000082709 00000 н 0000082842 00000 н 0000083021 00000 н 0000083202 00000 н 0000083333 00000 н 0000083515 00000 н 0000083697 00000 н 0000083864 00000 н 0000084015 00000 н 0000084220 00000 н 0000084407 00000 н 0000084570 00000 н 0000084775 00000 н 0000084963 00000 н 0000085126 00000 н 0000085291 00000 н 0000085472 00000 н 0000085663 00000 н 0000085843 00000 н 0000085981 00000 н 0000086135 00000 н 0000086304 00000 н 0000086454 00000 н 0000086602 00000 н 0000086787 00000 н 0000086947 00000 н 0000087141 00000 н 0000087348 00000 н 0000087478 00000 н 0000087634 00000 н 0000087846 00000 н 0000087988 00000 н 0000088167 00000 н 0000088307 00000 н 0000088442 00000 н 0000088598 00000 н 0000088750 00000 н 0000088902 00000 н 0000089034 00000 н 0000089188 00000 н 0000089376 00000 н 0000089544 00000 н 0000089710 00000 н 0000089884 00000 н 00000
00000 н 00000
00000 н 00000
00000 н 00000SEC.gov | Порог частоты запросов превысил
Чтобы обеспечить равный доступ для всех пользователей, SEC оставляет за собой право ограничивать запросы, исходящие от необъявленных автоматических инструментов. Ваш запрос был идентифицирован как часть сети автоматизированных инструментов, выходящих за рамки приемлемой политики, и будет управляться до тех пор, пока не будут предприняты действия по объявлению вашего трафика.
Пожалуйста, заявите о своем трафике, обновив свой пользовательский агент, включив в него информацию о компании.
Чтобы ознакомиться с рекомендациями по эффективной загрузке информации с SEC.gov, включая последние документы EDGAR, посетите сайт sec.gov/developer. Вы также можете подписаться на получение по электронной почте обновлений программы открытых данных SEC, включая передовые методы, которые делают загрузку данных более эффективной, и улучшения SEC.gov, которые могут повлиять на процессы загрузки по сценарию. Для получения дополнительной информации обращайтесь по адресу [email protected].
Для получения дополнительной информации см. Политику конфиденциальности и безопасности веб-сайта SEC.Благодарим вас за интерес, проявленный к Комиссии по ценным бумагам и биржам США.
Идентификатор ссылки: 0.67fd733e.1642658302.375fdc0b
Дополнительная информация
Политика безопасности Интернета
Используя этот сайт, вы соглашаетесь на мониторинг и аудит безопасности. В целях безопасности и для обеспечения того, чтобы общедоступные услуги оставались доступными для пользователей, эта правительственная компьютерная система использует программы для мониторинга сетевого трафика для выявления несанкционированных попыток загрузить или изменить информацию или иным образом нанести ущерб, включая попытки отказать в обслуживании пользователям.
Несанкционированные попытки загрузки информации и/или изменения информации в любой части этого сайта строго запрещены и подлежат судебному преследованию в соответствии с Законом о компьютерном мошенничестве и злоупотреблениях от 1986 г. и Законом о защите национальной информационной инфраструктуры от 1996 г. (см. Раздел 18 USC §§ 1001 и 1030).
Чтобы гарантировать, что наш веб-сайт хорошо работает для всех пользователей, SEC отслеживает частоту запросов на контент SEC.gov, чтобы гарантировать, что автоматический поиск не повлияет на возможность других получить доступ к SEC.содержание правительства. Мы оставляем за собой право блокировать IP-адреса, отправляющие чрезмерные запросы. Текущие правила ограничивают количество пользователей до 10 запросов в секунду, независимо от количества компьютеров, используемых для отправки запросов.
Если пользователь или приложение отправляет более 10 запросов в секунду, дальнейшие запросы с IP-адресов могут быть ограничены на короткий период. Как только количество запросов упадет ниже порогового значения на 10 минут, пользователь может возобновить доступ к контенту в SEC.правительство Эта практика SEC предназначена для ограничения чрезмерных автоматических поисков на SEC.gov и не предназначена и не ожидается, что она повлияет на отдельных лиц, просматривающих веб-сайт SEC.gov.
Обратите внимание, что эта политика может измениться, поскольку SEC управляет SEC.gov, чтобы обеспечить эффективную работу веб-сайта и его доступность для всех пользователей.
Примечание: Мы не предлагаем техническую поддержку для разработки или отладки процессов загрузки по сценарию.
границ | Терапия анти-PD-1 и анти-CTLA-4 при раке: механизмы действия, эффективность и ограничения
Введение
В последние годы произошел резкий скачок в разработке и внедрении противораковой иммунотерапии.Одобрение антител против цитотоксического белка 4, ассоциированного с Т-лимфоцитами (CTLA-4), и антител против белка программируемой гибели клеток 1 (PD-1) для использования человеком уже привело к значительному улучшению результатов лечения различных видов рака, особенно меланомы. . В отличие от радио- и химиотерапии, целью которых является непосредственное вмешательство в рост и выживание опухолевых клеток, иммунотерапия воздействует на опухоль косвенно, усиливая противоопухолевый иммунный ответ, который спонтанно возникает у многих пациентов.
Рак уклоняется от иммунного ответа и подавляет его
Чтобы понять способы действия ингибиторов иммунных контрольных точек, важно понять динамическое взаимодействие между раком и иммунной системой в ходе болезни.
Раковые клетки генетически нестабильны, что способствует их неконтролируемой пролиферации и экспрессии антигенов, распознаваемых иммунной системой. Эти антигены включают в себя нормальные белки, сверхэкспрессируемые раковыми клетками, и новые белки, которые образуются в результате мутаций и перестройки генов (1). Цитотоксические CD8 + Т-клетки представляют собой иммунные клетки, которые особенно эффективны в опосредовании противоопухолевого иммунного ответа. Эти клетки могут научиться распознавать опухолеспецифические антигены, представленные на молекулах главного комплекса гистосовместимости (MHC) класса I, и тем самым осуществлять целенаправленное уничтожение опухолевых клеток.CD8 + Т-клетки становятся лицензированными эффекторными клетками после соответствующей стимуляции антигенпрезентирующими клетками, которые собрали антигены в месте опухоли. Помимо антигенных пептидов, встроенных в молекулы MHC, антигенпрезентирующие клетки должны обеспечивать костимулирующие сигналы через поверхностные рецепторы (такие как CD28) и цитокины [такие как интерлейкин (IL)-12] для эффективной стимуляции Т-клеток (2).
Опухолевые клетки применяют различные механизмы, чтобы избежать иммунного распознавания и иммуноопосредованного разрушения.Часто считается, что установленные опухоли возникают в результате отбора клонов, способных ускользать от иммунной системы, процесс, известный как иммуноредактирование (3). Опухолевые клетки могут уклоняться от распознавания иммунной системой непосредственно за счет подавления функций, которые делают их уязвимыми, таких как опухолевые антигены или MHC класса I (4–6). С другой стороны, опухоли могут уклоняться от иммунного ответа, используя механизмы отрицательной обратной связи, которые организм выработал для предотвращения иммунопатологии. К ним относятся ингибирующие цитокины, такие как IL-10 и фактор роста опухоли (TGF)-β, ингибирующие типы клеток, такие как регуляторные Т-клетки (Treg), регуляторные В-клетки (Breg) и супрессорные клетки миелоидного происхождения (MDSC), модуляторы метаболизма. такие как индоламин-2,3-диоксигеназа (IDO) и ингибирующие рецепторы, такие как PD-1 и CTLA-4 (7, 8).
Иммунное истощение способствует иммунной дисфункции при раке
Ингибирующие рецепторы, также известные как иммунные контрольные точки, и их лиганды можно найти в клетках широкого спектра типов. Они необходимы для центральной и периферической толерантности, поскольку они противодействуют одновременной активации передачи сигналов через костимулирующие молекулы. Ингибирующие рецепторы могут действовать как во время иммунной активации, так и во время продолжающихся иммунных ответов. В частности, известно, что во время хронического воспаления Т-клетки истощаются и активируют широкий спектр неповторяющихся ингибирующих рецепторов, ограничивающих их эффективность, таких как PD-1, CTLA-4, Т-клеточный иммуноглобулин и муцин-домен, содержащий 3 (TIM-3), ген 3 активации лимфоцитов (LAG-3) или Т-клеточный иммунорецептор с доменами Ig и ITIM (TIGIT) [см. Таблицу 1 (9–11)].Первоначально описанный в контексте хронических вирусных инфекций, когда хозяину не удается избавиться от патогена, теперь очевидно, что истощение Т-клеток также может возникать при раке (12, 13). Считается, что в этих условиях постоянная высокая антигенная нагрузка приводит к тому, что Т-клетки активируют ингибирующие рецепторы, передача сигналов которых впоследствии приводит к прогрессирующей потере пролиферативного потенциала и эффекторных функций, а в некоторых случаях к их делеции (14).
Таблица 1 .Обзор рецепторов поверхности Т-клеток, связанных с иммунным торможением и дисфункцией.
Таким образом, истощение является как физиологическим механизмом, предназначенным для ограничения иммунопатологии во время персистирующей инфекции, так и основным препятствием для противоопухолевого иммунного ответа (17). Следует отметить, что экспрессия ингибиторных маркеров не всегда является признаком иммунного истощения, поскольку рецепторы могут экспрессироваться индивидуально во время обычных иммунных ответов (18).
Рецептор иммунной контрольной точки CTLA-4
Блокирующее антитело против CTLA-4 ипилимумаб было первым ингибитором контрольных точек иммунного ответа, который был протестирован и одобрен для лечения онкологических больных (19, 20).CTLA-4 (CD152) является членом семейства B7/CD28, который ингибирует функции Т-клеток. Он конститутивно экспрессируется Treg, но также может активироваться другими субпопуляциями Т-клеток, особенно Т-клетками CD4 + , при активации (21). Истощенные Т-клетки также часто характеризуются экспрессией CTLA-4 среди других ингибирующих рецепторов. CTLA-4 в основном локализуется во внутриклеточных везикулах и лишь временно экспрессируется при активации в иммунологическом синапсе, после чего быстро подвергается эндоцитозу (22).
CTLA-4 опосредует иммуносупрессию, опосредованно уменьшая передачу сигналов через костимулирующий рецептор CD28. Хотя оба рецептора связывают CD80 и CD86, CTLA-4 делает это с гораздо большей аффинностью, эффективно превосходя CD28 (23). CTLA-4 может также удалять CD80 и CD86 (включая их цитоплазматические домены) с клеточной поверхности антигенпрезентирующих клеток посредством трансэндоцитоза (24), тем самым снижая доступность этих стимулирующих рецепторов для других CD28-экспрессирующих Т-клеток.Действительно, этот процесс является важным механизмом, с помощью которого Treg опосредуют иммунную супрессию на клетках-свидетелях (25).
Ограничивая CD28-опосредованную передачу сигналов во время презентации антигена, CTLA-4 повышает порог активации Т-клеток, снижая иммунный ответ на слабые антигены, такие как собственные и опухолевые антигены. Центральная роль, которую CTLA-4 играет в иммунологической толерантности, иллюстрируется экспериментами на мышах, у которых отсутствует ген CTLA-4 глобально или конкретно в компартменте Forkhead box P3 (FoxP3) + Treg.У этих животных развиваются лимфопролиферативные заболевания, и они погибают в молодом возрасте (25, 26). Точно так же полиморфизмы в гене CTLA-4 связаны с аутоиммунными заболеваниями у людей (27). Было показано, что передача сигналов CTLA-4 ослабляет иммунный ответ против инфекций и опухолевых клеток (28, 29).
Рецептор иммунной контрольной точки PD-1
Поверхностный рецептор PD-1 (CD279) был впервые обнаружен на мышиной Т-клеточной гибридоме, и считалось, что он участвует в гибели клеток (30). Однако с тех пор стало ясно, что PD-1, который гомологичен CD28, в первую очередь участвует в ингибирующей иммунной передаче сигналов и является важным регулятором адаптивных иммунных ответов (31).Как у людей, так и у мышей некоторые популяции Т-клеток конститутивно экспрессируют PD-1; одним из примеров являются фолликулярные хелперные Т-клетки (32). Хотя большинство циркулирующих Т-клеток не экспрессируют рецептор, их можно индуцировать для этого при стимуляции с помощью комплекса Т-клеточного рецептора (TCR) или воздействия цитокинов, таких как IL-2, IL-7, IL-15, IL-1. 21 и трансформирующий фактор роста (TGF)-β (33, 34). Другие типы клеток, такие как В-клетки, миелоидные дендритные клетки, тучные клетки и клетки Лангерганса, также могут экспрессировать PD-1, который может регулировать свои собственные функции и функции клеток-свидетелей в патофизиологических условиях (35–38).PD-1 имеет два лиганда: PD-L1 (B7-h2; CD274) и PD-L2 (B7-DC; CD273). Оба могут быть обнаружены на поверхности антигенпрезентирующих клеток (таких как дендритные клетки, макрофаги и моноциты), но в остальном они по-разному экспрессируются в различных нелимфоидных тканях (39, 40). Интерферон (IFN)-γ является основным триггером, который, как известно, вызывает активацию PD-L1 и PD-L2 (41).
PD-1 несет мотив ингибирования на основе тирозина иммунорецептора (ITIM) и мотив переключения на основе тирозина иммунорецептора (ITSM) на своем внутриклеточном хвосте.События внутриклеточной передачи сигналов, инициированные при вовлечении PD-1, лучше всего описаны в Т-клетках и проиллюстрированы на рисунке 1. В этих клетках привлечение PD-1 вызывает фосфорилирование остатков тирозина, запуская внутриклеточный сигнальный каскад, который опосредует дефосфорилирование TCR. проксимальные сигнальные компоненты (9, 42–44). Среди них недавно было обнаружено, что CD28 является основной мишенью (45). В присутствии стимуляции TCR CD28 обеспечивает критические сигналы, важные для активации Т-клеток.Вмешиваясь в раннюю передачу сигналов TCR/CD28 и связанную с ними IL-2-зависимую положительную обратную связь, передача сигналов PD-1, таким образом, приводит к снижению продукции цитокинов [таких как IL-2, IFN-γ и фактор некроза опухоли (TNF)-α], прогрессирование клеточного цикла и экспрессия гена Bcl-xL, способствующая выживанию, а также снижение экспрессии факторов транскрипции, участвующих в эффекторных функциях, таких как T-bet и Eomes (42, 43, 46, 47). Таким образом, активность PD-1 актуальна только при одновременной активации Т-клеток, так как ее сигнальная трансдукция может вступать в силу только во время TCR-зависимой передачи сигналов (39, 41, 48).Детали передачи сигналов PD-1 в других типах клеток, несущих этот рецептор, таких как В-клетки, еще предстоит выяснить.
Рисунок 1 . Белок запрограммированной гибели клеток 1 (PD-1) опосредовал внутриклеточные сигнальные события во время активации Т-клеток. (1) При активации Т-клеток внеклеточные рецепторы PD-1, CD28 и комплекс Т-клеточных рецепторов (TCR) (включая CD4 или CD8) связывают свои лиганды PD-L1 или PD-L2, CD80 или CD86 и главный комплекс гистосовместимости (MHC) класса I или II соответственно.Это сближает все рецепторы друг с другом в иммунологическом синапсе и позволяет им взаимодействовать друг с другом. (2) Киназа Src Lck (P56Lck), которая связана с внутриклеточным хвостом CD4 и CD8, теперь может фосфорилировать остатки тирозина на внутриклеточных хвостах PD-1 и CD28, а также цепь CD3ζ TCR /CD3 комплекс. (3a) Фосфорилирование мотива переключателя на основе тирозина иммунорецептора (ITSM) на внутриклеточном хвосте PD-1 позволяет рекрутировать фосфатазу 2, содержащую домен гомологии Src 2 (SHP-2), что приводит к активации Фосфатазная активность SHP-2.SHP-1 также может связываться с PD-1, но в меньшей степени, чем SHP-2. (3b) Одновременно фосфорилированный хвост CD28 теперь способен рекрутировать PI-3K и Grb2 среди других сигнальных молекул. (4) За счет непосредственной близости к иммунологическому синапсу SHP-2, связанный с PD-1, может дефосфорилировать цитоплазматический хвост CD28 и, в меньшей степени, хвост цепи CD3ζ, тем самым предотвращая рекрутирование дальнейших нижестоящих сигнальных молекул, связанных с этими молекулами.SHP-2 может также дефосфорилировать PD-1, вызывая ауторегуляцию этого ингибирующего пути. (5) CD28 обеспечивает критические сигналы наряду со стимуляцией TCR, поэтому отмена связывания PI3K и Grb2 с этим рецептором приводит к снижению передачи сигналов в путях, важных для продукции IL-2, выживания, пролиферации и некоторых эффекторных функций. В отсутствие своих лигандов PD-1 не рекрутируется в иммунный синапс и, следовательно, не может мешать передаче сигналов активации. (6) Ингибирующий рецептор CTLA-4 главным образом косвенно ограничивает передачу сигналов CD28 за счет снижения доступности CD80 и CD86, с которыми он связывается с гораздо более высокой аффинностью, чем костимулирующий рецептор CD28.Источники (43–45).
В целом, PD-1 имеет решающее значение для поддержания периферической толерантности и сдерживания иммунных реакций во избежание иммунопатологии. Мыши с дефицитом рецептора изначально кажутся здоровыми, но с возрастом у них развиваются аутоиммунные заболевания, такие как волчаночноподобный пролиферативный гломерулонефрит и артрит, а также обострение воспаления во время инфекций (18, 31, 49, 50). Люди с генетическим полиморфизмом в локусе PD-1 также имеют повышенную вероятность развития различных аутоиммунных заболеваний (51, 52).
CTLA-4, PD-1 и их лиганды при раке
CTLA-4 может экспрессироваться в опухолевых поражениях на инфильтрирующих Treg или истощенных обычных Т-клетках, а также в самих опухолевых клетках (53, 54). Несмотря на иммуносупрессивную роль CTLA-4, его связь с прогнозом заболевания не ясна; однако следует отметить, что лишь в нескольких исследованиях описано прогностическое значение уровней CTLA-4 в очаге опухоли. До сих пор экспрессия CTLA-4 в опухолях была связана со снижением выживаемости при карциноме носоглотки (54) и повышением выживаемости при немелкоклеточном раке легкого (53).
PD-1 может кратковременно активироваться во время стимуляции и конститутивно во время хронической иммунной активации (17). Ингибирующий рецептор был обнаружен как на циркулирующих опухолеспецифических Т-клетках, так и на инфильтрирующих опухоль лимфоцитах, где он был связан со снижением функции Т-клеток у людей и мышей (13, 29, 55–57). Другие типы клеток также могут активировать PD-1 при опухолевых поражениях. PD-1-положительные дендритные клетки, например, были идентифицированы при гепатоцеллюлярной карциноме, где они проявляли сниженную способность стимулировать Т-клетки (37).В другом исследовании идентифицирована популяция инфильтрирующих опухоль регуляторных В-клеток, экспрессирующих PD-1, которые продуцируют IL-10; более высокие доли этих клеток коррелировали с худшим исходом заболевания у пациентов с гепатоцеллюлярной карциномой (58). Также недавно было показано, что ассоциированные с опухолью макрофаги экспрессируют PD-1 как у мышей, так и у людей с колоректальным раком и нарушают фагоцитоз макрофагов (59).
Как раковые клетки, так и проникающие в опухоль иммунные клетки (такие как макрофаги) могут экспрессировать PD-L1 и активировать его в ответ на IFN-γ (60).Таким образом, экспрессия PD-L1 может свидетельствовать об активных противоопухолевых иммунных реакциях, а также может активно способствовать местной иммуносупрессии. Таким образом, взаимосвязь между экспрессией PD-1 или PD-L1 в месте опухоли и исходом заболевания неодинакова для всех типов опухолей и пациентов. Высокий уровень PD-1 и/или PD-L1 может коррелировать с плохим прогнозом при некоторых видах рака (включая меланому, почечно-клеточную карциному, рак пищевода, желудка и яичников) и с улучшенным прогнозом при других (таких как ангиосаркома и рак желудка) (55). , 60–65).
Эффективность и механизм действия ингибиторов контрольных точек
Ингибиторы контрольных точек CTLA-4 и PD-1 привели к увеличению выживаемости пациентов в ряде исследований, включая исследования меланомы, почечно-клеточного рака, плоскоклеточного рака и немелкоклеточного рака легкого, по сравнению с традиционной химиотерапией ( обобщены в таблице 2). При меланоме лечение анти-PD-1 было более эффективным у пациентов с небольшими опухолями (66). Прямое сравнение между двумя ингибиторами контрольных точек в клиническом испытании фазы III показало лучший ответ (44%) и показатели выживаемости (6.9-месячная выживаемость без прогрессирования) среди пациентов, получавших ниволумаб с антителом к PD-1, чем среди пациентов, получавших ипилимумаб с антителом к CTLA-4 (19% и 2,8 месяца). Комбинированное введение ниволумаба и ипилимумаба привело к еще более высокой частоте ответа (58%) и выживаемости (11,5 месяцев) (67).
Таблица 2 . Результаты лечения клинических испытаний ингибиторов иммунных контрольных точек при различных типах рака.
Как CTLA-4, так и PD-1 действуют независимо как тормоза на CD3/CD28-зависимую передачу сигналов, что свидетельствует о том, что лежащие в основе иммунные реакции необходимы для того, чтобы лечение ингибитором контрольной точки вступило в силу (66).Действительно, как упоминалось в предыдущем разделе, блокада как PD-1, так и CTLA-4 более эффективна при опухолях, инфильтрированных Т-клетками или имеющих высокую частоту мутаций и, следовательно, более иммуногенных до лечения (86–88).
Прямые иммунологические последствия лечения анти-PD-1 и анти-CTLA-4 в основном исследовались на Т-клетках (рис. 2). Считается, что блокада CTLA-4, скорее всего, влияет на стадию активации Т-клеток в дренирующих лимфатических узлах, когда Treg, экспрессирующие CTLA-4, удаляют CD80/CD86 с поверхности антигенпрезентирующих клеток, тем самым снижая их способность эффективно стимулировать опухолеспецифические Т-клетки (24).Блокада CTLA-4 может также воздействовать на место опухоли, поскольку истощенные Т-клетки, экспрессирующие CTLA-4, и Treg могут накапливаться в микроокружении опухоли (29, 53). Экспрессирующие PD-1 инфильтрирующие опухоль Т-клетки могут быть инактивированы PD-L1 на поверхности опухолевых клеток или других инфильтрирующих иммунных клеток, и поэтому считается, что блокирующие антитела, нацеленные на передачу сигналов PD-1, в основном влияют на эффекторную стадию иммунного ответа. (13, 55–57). Поскольку другие типы клеток (такие как дендритные клетки и В-клетки) также могут находиться под влиянием передачи сигналов PD-1, ингибирование пути PD-1/PD-L1 также может иметь независимые от Т-клеток эффекты, которые влияют на иммунные ответы во время контрольной точки. Ингибиторную терапию еще предстоит выяснить (36, 58).
Рисунок 2 . Роль белка запрограммированной гибели клеток 1 (PD-1) и белка 4, ассоциированного с Т-лимфоцитами (CTLA-4), в прайминговой и эффекторной фазах противоопухолевого иммунного ответа. Для примирования Т-клеток дендритные клетки (ДК) берут образец антигена в месте опухоли и транспортируют его в дренирующие лимфатические узлы, где они представляют антигены на своих молекулах главного комплекса гистосовместимости (МНС) Т-клеткам. Т-клетки активируются, если их рецепторы Т-клеток распознают и связывают антиген на комплексах MHC, а их костимулирующие рецепторы CD28 связывают CD80 и CD86 на DC.Повышающая регуляция CTLA-4 на Т-клетках или сторонних Treg может мешать сигналу CD28, поскольку первый рецептор связывает CD80 и CD86 с более высокой аффинностью. После активации Т-клетки мигрируют к месту опухоли, чтобы убить злокачественные клетки. Опухоли или клетки-свидетели, такие как макрофаги, могут, однако, активировать PD-L1 и, следовательно, препятствовать функции Т-клеток, индуцируя ингибирующую внутриклеточную передачу сигналов. Таким образом, блокирующее антитело против CTLA-4 может восстанавливать примирование Т-клеток в лимфатических узлах, а блокада передачи сигналов PD-1 может активировать эффекторную функцию Т-клеток в месте опухоли.Кроме того, другие типы клеток, такие как клетки Breg и DC в микроокружении опухоли, могут экспрессировать PD-1 и, следовательно, подвергаться блокаде PD-1. Блокада PD-1 и CTLA-4 также может влиять на профили Т-хелперов напрямую или путем воздействия на микробиоту.
Иммунный ответ типа I, который включает продукцию IFN-γ и функции цитотоксических Т-клеток, важен для эффективного противоопухолевого иммунного ответа и связан с лучшим ответом на лечение анти-CTLA-4 и анти-PD-1. Действительно, мышиные модели показали, что локальная активация IFN-γ необходима для анти-PD-1-опосредованной регрессии опухоли (89).Точно так же IFN-γ и компонент цитотоксических гранул гранзим B были повышены в регрессирующих поражениях у пациентов с меланомой после лечения анти-PD-1 (90). Опухоли у пациентов, получавших анти-PD-1, которые сначала ответили, а затем рецидивировали, показали мутации, которые вызвали последующую потерю поверхностной экспрессии MHC класса I (чтобы избежать распознавания цитотоксическими Т-клетками) или элементов ответа IFN-γ (6). Т-клетки Th9 CD4 + также предполагают свою роль в соответствии с недавним исследованием, в котором обнаружено значительное увеличение частоты клеток Th9 у пациентов, отвечающих на лечение анти-PD-1 (91, 92).
Может показаться заманчивым предположить, что ингибиторы иммунных контрольных точек специфически усиливают функцию Т-клеток, принадлежащих к эффекторному компартменту памяти, поскольку эти клетки легко экспрессируют цитотоксические молекулы, такие как перфорин и гранзим B. Однако в этих клетках отсутствует костимулирующий рецептор CD28. благодаря чему как PD-1, так и CTLA-4 ингибируют функцию Т-клеток (93). Два недавних исследования показали, что именно CD28-экспрессирующие клетки, а не уже терминально дифференцированные эффекторные клетки, реагируют на блокаду PD-1 пролиферативным всплеском и дифференцировкой (94, 95).
Характеристики самой опухоли также могут влиять на эффективность ингибитора иммунных контрольных точек. Мутационная нагрузка опухолевых клеток может повышать их антигенность, но также может усиливать их способность уклоняться от иммунного ответа, вызванного лечением. Действительно, недавнее исследование выявило сигнатуру гена меланомы, связанную с врожденной резистентностью к анти-PD-1, которая включала активацию генов, связанных с ангиогенезом, заживлением ран, мезенхимальным переходом, клеточной адгезией и ремоделированием внеклеточного матрикса (96).
Комменсальные бактерии также могут влиять на эффективность ингибиторов контрольных точек иммунитета. Было обнаружено, что лечение анти-CTLA-4 неэффективно у мышей, выращенных в стерильных условиях, и вызывает изменение кишечной флоры у мышей, выращенных обычным образом. Дальнейшие эксперименты показали, что присутствие определенных бактериальных штаммов, в частности Bacteroides fragilis , способствовало поляризации Th2 у животных и было связано с улучшенным противоопухолевым иммунным ответом (97).Важно отметить, что лечение антибиотиками также было связано со снижением ответа на лечение анти-PD-1/PD-L1 у онкологических больных, возможно, за счет изменения нормальной кишечной флоры. Вместо этого хороший ответ на лечение у пациентов был связан с присутствием комменсала Akkermansia muciniphila , который также улучшал ответ на лечение анти-PD-1 у мышей, позволяя увеличить привлечение CCR9 + CXCR3 + CD4 + T-лимфоцитов в опухоль (98). .
Нежелательные явления, связанные с лечением, и их лечение
PD-1 и CTLA-4 предотвращают аутоиммунитет и ограничивают иммунную активацию, чтобы предотвратить повреждение посторонними лицами в физиологических условиях.Таким образом, ингибирование этих рецепторов с помощью терапевтических антител для лечения рака связано с широким спектром побочных эффектов, которые напоминают аутоиммунные реакции. Частота серьезных побочных эффектов сильно различается в зависимости от исследования и лечения (см. Таблицу 2). Клинические испытания, в которых непосредственно сравнивали различные типы ингибиторов иммунных контрольных точек и их комбинации, показали, что большее количество пациентов испытывали побочные эффекты при лечении анти-CTLA-4 (27,3%) по сравнению с анти-PD-1 (16,3%). Еще больше пациентов пострадали при лечении комбинацией обоих (55%) (67).
Почти все пациенты, получающие ингибиторы контрольных точек иммунитета, испытывают легкие побочные эффекты, такие как диарея, утомляемость, зуд, сыпь, тошнота и снижение аппетита. Тяжелые побочные реакции включают тяжелую диарею, колит, повышение уровня аланинаминотрансферазы, воспалительный пневмонит и интерстициальный нефрит (67, 73, 99). Также были сообщения об обострении ранее существовавших аутоиммунных заболеваний, таких как псориаз (91, 92, 100), или развитии новых, таких как сахарный диабет 1 типа (101).При особо тяжелых побочных эффектах может потребоваться прекращение лечения, хотя эти пациенты могут по-прежнему реагировать после этого (102). Интересно, что некоторые аутоиммунные реакции, связанные с лечением, такие как сыпь и витилиго, коррелируют с лучшим прогнозом заболевания (103), что свидетельствует о совпадении аутоиммунных и противоопухолевых иммунных реакций.
Биомаркеры эффективности лечения ANTI-PD-1/CTLA-4
Биомаркеры необходимы как до, так и во время лечения, чтобы выявить пациентов, которые наиболее или наименее вероятно ответят на лечение ингибиторами контрольных точек иммунитета, чтобы уменьшить неадекватное воздействие лекарств.Ответ на лечение определяется как уменьшение размера опухоли в ходе курса лечения. Ряд факторов, связанных с прогнозом заболевания у нелеченных пациентов, также связан с частотой ответа ингибиторов иммунных контрольных точек (таблица 3). Например, пациенты с меньшими опухолями или низким уровнем лактатдегидрогеназы (ЛДГ) в сыворотке крови на исходном уровне имеют лучший прогноз и также с большей вероятностью реагируют на лечение анти-PD-1 (66). Снижение уровня ЛДГ после лечения также связано с улучшением ответа (104).Циркулирующая опухолевая ДНК (цДНК), которая содержит мутации, связанные с меланомой, и может высвобождаться мертвыми опухолевыми клетками, может быть обнаружена в сыворотке крови некоторых пациентов. Уровни CtDNA сильно коррелируют с опухолевой массой и прогрессированием (105, 106). Недавнее исследование пациентов с меланомой на поздних стадиях, получавших анти-PD-1 (отдельно или в комбинации с анти-CTLA-4), показало высокую частоту ответа на лечение у лиц, которые были отрицательными по цДНК до или после лечения (107). привлекательный биомаркер до и во время лечения иммунных контрольных точек.
Таблица 3 . Биомаркеры, связанные с благоприятным ответом на ингибиторы иммунных контрольных точек.
Для лечения анти-PD-1 экспрессия PD-L1 в микроокружении опухоли была очевидным кандидатом в биомаркеры. Хотя экспрессия PD-L1 на опухолевых клетках коррелировала с эффективностью лечения у пациентов с меланомой (67, 108), этого не было у пациентов с плоскоклеточным раком, немелкоклеточным раком легкого и карциномой из клеток Меркеля (70, 72, 74). Интересно, что одно исследование, оценивающее роль PD-L1 как в раковых клетках, так и в опухолевых иммунных клетках, показало, что только в последнем контексте эффективность лечения анти-PD-L1 коррелировала с экспрессией PD-L1 (77).
Присутствие неоантигенов на мутировавших опухолевых клетках повышает противоопухолевую иммуногенность и повышает эффективность лечения. Таким образом, высокая генетическая разница между опухолевыми клетками и клетками-хозяевами является показателем эффективности лечения ингибиторами контрольных точек. Это было особенно заметно у пациентов с меланомой, получавших анти-CTLA-4, в опухолях которых обнаруживались неоантигены (87), а также у пациентов, получавших анти-PD-1, с колоректальным раком или немелкоклеточным раком легкого, у которых был дефицит репарации несоответствия. или имели высокую частоту мутаций, соответственно (78, 86).Хотя общее мутационное бремя связано с улучшенным ответом на лечение анти-PD-1, сниженный ответ был обнаружен у пациентов с меланомой, опухоли которых демонстрировали сигнатуру гена IPRES (96). Презентация антигена хозяином также может играть роль во время лечения анти-PD-1, поскольку у пациентов с HLA-A*26 вероятность ответа была более чем в два раза выше, чем у пациентов, отрицательных по аллелю (111).
Другие иммунологические факторы до лечения, связанные с улучшенным ответом на лечение, включают высокий уровень эозинофилов и лимфоцитов в крови, обилие CD8 + Т-клеток, инфильтрирующих опухоль или присутствующих на краю опухоли, и повышенные уровни TGF-β в сыворотке у пациентов с меланомой, получавших лечение. с анти-PD-1 (88, 91, 92, 109).Повышенные уровни экспрессии генов Th2 и CTLA-4 (но не FoxP3) также были отмечены у пациентов с ответом на лечение с различными солидными опухолями (включая меланому), получавших анти-PD-L1 (77).
Ряд иммунологических наблюдений после лечения также был связан с улучшением ответа иммунных ингибиторов контрольных точек. Например, пациенты, которые с большей вероятностью реагируют на лечение анти-CTLA-4, имели повышенное количество Т-клеток, экспрессирующих индуцибельную костимулирующую молекулу (ICOS), и более низкое соотношение нейтрофилов к лимфоцитам (110).Увеличение пролиферации Т-клеток CD8 + в опухолевом очаге и повышенная частота Th9-клеток в кровотоке пациентов также были связаны с ответом на лечение (88, 91, 92).
В совокупности многие из этих исследований показывают, что ингибиторы контрольных точек иммунного ответа наиболее эффективны у пациентов, у которых уже до начала терапии проявляются противоопухолевые иммунные процессы. Однако не все биомаркеры, перечисленные здесь, могут быть одинаково эффективны, и пациенты могут по-прежнему реагировать на лечение, несмотря на противоположные прогнозы, основанные на биомаркерах.Кроме того, доступ к опухолевой ткани может быть затруднен у многих пациентов, особенно после лечения, и поэтому менее инвазивные «жидкие биопсии» на основе крови могут быть более подходящими. Важно отметить, что было показано, что исследование нескольких биомаркеров в сочетании может улучшить прогноз лечения (109). Хотя недавно открытая цтДНК кажется особенно многообещающим кандидатом в биомаркеры, необходимы дополнительные исследования для выявления более эффективных биомаркеров или комбинаций биомаркеров, чтобы разработать наиболее подходящую стратегию лечения для каждого пациента.
Ограничения ингибиторов иммунных контрольных точек
Хотя лечение ингибиторами контрольных точек иммунного ответа может быть эффективным на начальном этапе, у многих пациентов в конечном итоге возникает рецидив и прогрессирование опухоли. Поэтому в ряде исследований была предпринята попытка понять механизмы, с помощью которых лечение анти-PD-1 и анти-CTLA-4 теряет свою эффективность.
Давление отбора, вызванное лечением ингибитором контрольной точки, может привести к возникновению опухолевых клеток, которые могут уклоняться от иммуноопосредованного распознавания и делеции по новым путям.Например, недавно было показано, что опухолевые клетки пациентов, рефрактерных к лечению анти-PD-1, приобрели мутации, делающие их менее восприимчивыми к опосредованному Т-клетками уничтожению за счет потери ответных элементов IFN-γ или MHC класса I (6) .
Лечение анти-PD-1 или анти-CTLA-4 может также вызвать активацию других ингибирующих рецепторов. Например, у пациентов с меланомой или раком предстательной железы наблюдалась активация ингибирующего рецептора V-домена Ig-супрессора активации Т-клеток (VISTA) на различных инфильтрирующих опухоль иммунных клетках после лечения анти-CTLA-4 (112).В другом исследовании была отмечена активация ингибирующего рецептора TIM-3 (но не VISTA) на поверхности Т-клеток у мышей, получавших анти-PD-1, с раком легкого, а также активация TIM-3 на Т-клетках у пациентов с аденокарциномой, резистентных к Лечение ПД-1 (113).
Совсем недавно исследование выявило еще один неожиданный механизм резистентности к анти-PD-1 терапии у мышей, при котором макрофаги, ассоциированные с опухолью, удаляли терапевтическое антитело с поверхности Т-клеток in vivo , тем самым снова делая их восприимчивыми к ингибирующей передаче сигналов через рецептор.Это явление может быть частично устранено введением агентов, блокирующих Fc-рецепторы, перед лечением (114). Таким образом, лучшее понимание механизмов, ограничивающих эффективность ингибиторов иммунных контрольных точек, позволит улучшить методы лечения в будущем.
Перспективы будущего: расширение репертуара лечения ингибиторами контрольных точек иммунитета
Блокаторы PD-1 и CTLA-4 не эффективны у всех пациентов, и даже у тех пациентов, которые изначально реагируют, могут возникнуть рецидивы, что подчеркивает необходимость улучшенных или альтернативных методов лечения.Были идентифицированы альтернативные ингибирующие рецепторы, которые также могут быть нацелены на противоопухолевую иммунную терапию. К ним относятся рецепторы TIM-3, LAG-3, TIGIT и B- и T-лимфоцит-ассоциированного белка (BTLA), связанные с истощением Т-клеток, а также VISTA, рецептор, обнаруженный на инфильтрирующих опухоль миелоидных клетках, ингибирование которого способствует противоопухолевые иммунные ответы в мышиных моделях и CD96, который, как было показано, ингибирует активность NK-клеток в мышиных моделях рака (115–117).
Комбинации ингибиторов контрольных точек иммунного ответа друг с другом или с другими видами лечения также изучаются.Действительно, комбинация лечения против CTLA-4 с лечением против PD-1 показала более высокую эффективность по сравнению с индивидуальным введением, но также была связана с усилением побочных эффектов. Фермент IDO, метаболизирующий триптофан, ингибирует функцию Т-клеток, а комбинация препаратов, блокирующих IDO, вместе с ингибиторами иммунных контрольных точек показала многообещающие результаты на мышах и в настоящее время также проходит клинические испытания на людях (105, 118). Макрофаги могут также влиять на противоопухолевый иммунитет или даже напрямую ограничивать терапевтические антитела (114).Поэтому их истощение с помощью ингибитора рецептора колониестимулирующего фактора-1 (CSF-1R) изучается в клинических испытаниях вместе с анти-PD-1 после того, как была показана эффективность на мышиной модели глиобластомы (119). Противоопухолевая функция Т-клеток, вызванная блокадой PD-1 у мышей, также может быть улучшена за счет целенаправленного увеличения функции митохондрий (120).
Поскольку ингибиторы иммунных контрольных точек работают, устраняя тормоза в иммунной системе, а не напрямую усиливая иммунную функцию, пациентам также может помочь комбинированная терапия, включающая иммуностимулирующие вещества.Модели меланомы у мышей, например, показали, что комбинация анти-CTLA-4 с цитокинами, такими как гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), или с агонистическими антителами, нацеленными на костимуляторные рецепторы, такие как CD40, усиливала отторжение опухоли в синергетическим образом (121, 122). Генетически модифицированный вирус простого герпеса talimogene laherparepvec предназначен для репликации в опухолевых клетках и высвобождения GM-CSF, тем самым привлекая иммунные клетки в среду опухоли. Вирус был протестирован в недавних клинических испытаниях в сочетании либо с CTLA-4, либо с PD-1 у пациентов с меланомой на поздних стадиях, что привело к увеличению частоты ответа на лечение по сравнению с одними ингибиторами иммунных контрольных точек (123, 124).
Даже модуляция кишечного микробиома может улучшить терапию на основе ингибиторов контрольных точек иммунитета. Введение только кишечных Bifidobacteria было связано со снижением роста опухоли в мышиной модели меланомы B16 за счет стимуляции опосредованных дендритными клетками ответов CD8 + T-клеток. Важно отметить, что введение этих бактерий также усиливало терапевтический эффект лечения анти-PD-1 у этих мышей (125). В аналогичном исследовании введение B. fragilis стерильным мышам, получавшим анти-CTLA-4, приводило к снижению роста опухоли, скорее всего, за счет благоприятного сдвига в сторону Th2-ответов (97).Исследования на людях также позволили связать присутствие фекальных A. muciniphila, Ruminococcaceae и Faecalibacterium с благоприятным исходом лечения анти-PD-1 (98, 126). получить пользу от надлежащего управления их кишечной флорой во время лечения ингибиторами иммунных контрольных точек.
Таким образом, в настоящее время изучается широкий спектр многообещающих новых направлений, хотя их клиническая эффективность еще предстоит подтвердить в текущих и будущих клинических испытаниях.
Заключительные замечания
Хотя терапия, направленная на PD-1 и CTLA-4, смогла увеличить среднюю продолжительность жизни больных раком, смертность среди пациентов с поздними стадиями остается высокой, что подчеркивает необходимость дальнейших инноваций в этой области. Как анти-PD-1, так и анти-CTLA-4 терапия, по-видимому, более эффективна у пациентов с уже существующим противоопухолевым иммунитетом, что позволяет предположить, что у пациентов без такого иммунитета эти препараты не способны опосредовать противоопухолевый иммунный ответ де ново .Однако по мере того, как наше понимание механизмов действия этих препаратов улучшается, открываются возможности для улучшения их использования не только за счет целенаправленного воздействия на тех пациентов, которые с наибольшей вероятностью реагируют с помощью соответствующих процедур скрининга биомаркеров, но и за счет сочетания используемых в настоящее время ингибиторов контрольных точек иммунного ответа с другие дополнительные препараты, чтобы помочь тем пациентам, которые не реагируют на текущие схемы лечения.
Вклад авторов
Этот обзор был подготовлен JAS и AO и критически отредактирован KK.
Заявление о конфликте интересов
АОполучил исследовательский грант от ONO PHARMACEUTICAL CO., LTD. и Бристол-Майерс Сквибб.
Финансирование
Эта работа была частично поддержана грантами на научные исследования от Министерства образования, культуры, спорта, науки и технологий и Министерства здравоохранения, труда и социального обеспечения Японии.
Сокращения
ctDNA, кольцевая ДНК опухоли; CTLA-4, белок 4, ассоциированный с Т-лимфоцитами; DCs, дендритные клетки; IPRES, врожденная устойчивость к анти-PD-1; ЛДГ, лактатдегидрогеназа; NK, естественные клетки-киллеры; PD-1, белок 1 запрограммированной гибели клеток; IDO, индоламин-2,3-диоксигеназа; ИЛ-12, интерлейкин 12; TGF-β, фактор роста опухоли-β; Tregs, регуляторные Т-клетки; MDSCs, супрессорные клетки миелоидного происхождения; VISTA, Ig-супрессор V-домена активации Т-клеток; ITIM, мотив ингибирования иммунорецептора на основе тирозина; ИФН-γ, интерферон; TIM-3, Т-клеточный иммуноглобулин и муцин-домен, содержащий-3; LAG-3, белок 3 активации лимфоцитов; TIGIT, Т-клеточный иммунорецептор с доменами Ig и ITIM; TNF-α, фактор некроза опухоли-α; ICOS, индуцибельная костимулирующая молекула; ИФН-γ, интерферон-γ; BTLA, белок, ассоциированный с В- и Т-лимфоцитами; CSF-1R, рецептор колониестимулирующего фактора-1; ГМ-КСФ, гранулоцитарно-макрофагальный колониестимулирующий фактор; Breg, регуляторная В-клетка.
Ссылки
1. Лоуренс М.С., Стоянов П., Полак П., Крюков Г.В., Цибульскис К., Сиваченко А. и соавт. Мутационная гетерогенность при раке и поиск новых генов, ассоциированных с раком. Природа (2013) 499(7457):214–8. дои: 10.1038/природа12213
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
2. Пеннок Н.Д., Уайт Дж.Т., Кросс Э.В., Чейни Э.Е., Тамбурини Б.А., Кедл Р.М. Реакция Т-клеток: наивная память и все, что между ними. Adv Physiol Educ (2013) 37(4):273–83.doi:10.1152/advan.00066.2013
Полнотекстовая перекрестная ссылка | Академия Google
4. Otsuka A, Dreier J, Cheng PF, Nägeli M, Lehmann H, Felderer L, et al. Ингибиторы пути Hedgehog способствуют адаптивным иммунным ответам при базально-клеточной карциноме. Clin Cancer Res (2015) 21(6):1289–97. doi:10.1158/1078-0432.CCR-14-2110
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
6. Zaretsky JM, Garcia-Diaz A, Shin DS, Escuin-Ordinas H, Hugo W, Hu-Lieskovan S, et al.Мутации, связанные с приобретенной резистентностью к блокаде PD-1 при меланоме. N Engl J Med (2016) 375(9):819–29. дои: 10.1056/NEJMoa1604958
Полнотекстовая перекрестная ссылка | Академия Google
7. Vinay DS, Ryan EP, Pawelec G, Talib WH, Stagg J, Elkord E, et al. Уклонение от иммунитета при раке: механистическая основа и терапевтические стратегии. Semin Cancer Biol (2015) 35 (Приложение): S185–98. doi:10.1016/j.semcancer.2015.03.004
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
9.Парри Р.В., Хемниц Дж.М., Фраувирт К.А., Ланфранко А.Р., Браунштейн И., Кобаяши С.В. и др. Рецепторы CTLA-4 и PD-1 ингибируют активацию Т-клеток по разным механизмам. Mol Cell Biol (2005) 25(21):9543–53. doi: 10.1128/MCB.25.21.9543-9553.2005
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
10. Blackburn SD, Shin H, Haining WN, Zou T, Workman CJ, Polley A, et al. Корегуляция истощения CD8+ Т-клеток множественными ингибирующими рецепторами при хронической вирусной инфекции. Nat Immunol (2009) 10(1):29–37. дои: 10.1038/ni.1679
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
11. Андерсон А.С., Джоллер Н., Кучроо В.К. Lag-3, Tim-3 и TIGIT: коингибирующие рецепторы со специализированными функциями в иммунной регуляции. Иммунитет (2016) 44(5):989–1004. doi:10.1016/j.immuni.2016.05.001
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
12. Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, Reddy S, et al.Экспрессия PD-1 на ВИЧ-специфических Т-клетках связана с истощением Т-клеток и прогрессированием заболевания. Природа (2006) 443(7109):350–4. дои: 10.1038/природа05115
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
13. Baitsch L, Baumgaertner P, Devêvre E, Raghav SK, Legat A, Barba L, et al. Истощение опухолеспецифических CD8+ Т-клеток в метастазах больных меланомой. J Clin Invest (2011) 121(6):2350–60. дои: 10.1172/JCI46102
Полнотекстовая перекрестная ссылка | Академия Google
16.Гарднер Д., Джеффри Л.Э., Сэнсом Д.М. Понимание пути CD28/CTLA-4 (CD152) и его значения для костимулирующей блокады. Am J Transplant (2014) 14 (9): 1985–91. дои: 10.1111/ajt.12834
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
17. Speiser DE, Utzschneider DT, Oberle SG, Münz C, Romero P, Zehn D. Дифференцировка Т-клеток при хронической инфекции и раке: функциональная адаптация или истощение? Nat Rev Immunol (2014) 14(11):768–74.дои: 10.1038/nri3740
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
18. Erickson JJ, Gilchuk P, Hastings AK, Tollefson SJ, Johnson M, Downing MB, et al. Вирусные острые инфекции нижних дыхательных путей повреждают CD8+ Т-клетки через PD-1. J Clin Invest (2012) 122(8):2967–82. дои: 10.1172/JCI62860
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
19. Hodi FS, Mihm MC, Soiffer RJ, Haluska FG, Butler M, Seiden MV, et al. Биологическая активность блокады цитотоксических Т-лимфоцитов, ассоциированных с антигеном 4, у ранее вакцинированных пациентов с метастатической меланомой и карциномой яичников. Proc Natl Acad Sci USA (2003) 100(8):4712–7. doi:10.1073/pnas.0830997100
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
20. Phan GQ, Yang JC, Sherry RM, Hwu P, Topalian SL, Schwartzentruber DJ и др. Регрессия рака и аутоиммунитет, вызванные блокадой антигена 4, ассоциированного с цитотоксическими Т-лимфоцитами, у пациентов с метастатической меланомой. Proc Natl Acad Sci U S A (2003) 100(14):8372–7. doi:10.1073/pnas.1533209100
Полнотекстовая перекрестная ссылка | Академия Google
21.Чан Д.В., Гибсон Х.М., Ауфьеро Б.М., Уилсон А.Дж., Хафнер М.С., Ми Q-S и др. Дифференциальная экспрессия CTLA-4 в CD4+ Т-клетках человека по сравнению с CD8+ Т-клетками связана с повышением уровня NFAT1 и ингибированием пролиферации CD4+. Genes Immun (2014) 15(1):25–32. doi:10.1038/gene.2013.57
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
22. Леунг Х.Т., Брэдшоу Дж., Кливленд Дж.С., Линсли П.С. Ассоциированная с цитотоксическими Т-лимфоцитами молекула-4, высокоавидный рецептор для CD80 и CD86, содержит мотив внутриклеточной локализации в своем цитоплазматическом хвосте. J Biol Chem (1995) 270(42):25107–14. дои: 10.1074/jbc.270.42.25107
Полнотекстовая перекрестная ссылка | Академия Google
24. Qureshi OS, Zheng Y, Nakamura K, Attridge K, Manzotti C, Schmidt EM, et al. Трансэндоцитоз CD80 и CD86: молекулярная основа внеклеточной функции CTLA-4. Наука (2011) 332(6029):600–3. doi:10.1126/наука.1202947
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
25. Wing K, Onishi Y, Prieto-Martin P, Yamaguchi T, Miyara M, Fehervari Z, et al.CTLA-4 контролирует регуляторную функцию Foxp3+ Т-клеток. Science (2008) 322(5899):271–5. doi:10.1126/наука.1160062
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
26. Waterhouse P, Penninger JM, Timms E, Wakeham A, Shahinian A, Lee KP, et al. Лимфопролиферативные заболевания с ранней летальностью у мышей с дефицитом Ctla-4. Science (1995) 270(5238):985–8. doi:10.1126/наука.270.5238.985
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
28.Накамото Н., Чо Х., Шакед А., Олтофф К., Валига М.Е., Камински М. и др. Синергическое обращение вспять внутрипеченочного истощения HCV-специфических CD8 Т-клеток за счет комбинированной блокады PD-1/CTLA-4. PLoS Pathog (2009) 5(2):e1000313. doi:10.1371/journal.ppat.1000313
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
29. Карран М.А., Монтальво В., Ягита Х., Эллисон Дж.П. Комбинированная блокада PD-1 и CTLA-4 увеличивает количество инфильтрирующих Т-клеток и уменьшает количество регуляторных Т-клеток и миелоидных клеток в опухолях меланомы B16. Proc Natl Acad Sci U S A (2010) 107(9):4275–80. doi:10.1073/pnas.0
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
30. Ishida Y, Agata Y, Shibahara K, Honjo T. Индуцированная экспрессия PD-1, нового члена суперсемейства генов иммуноглобулинов, при запрограммированной гибели клеток. EMBO J (1992) 11(11):3887.
Резюме PubMed | Академия Google
31. Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Развитие волчаночноподобных аутоиммунных заболеваний путем нарушения гена PD-1, кодирующего иммунорецептор, несущий мотив ITIM. Иммунитет (1999) 11(2):141–51. дои: 10.1016/S1074-7613(00)80089-8
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
32. Сейдж П.Т., Франсиско Л.М., Карман К.В., Шарп А.Х. Рецептор PD-1 контролирует фолликулярные регуляторные Т-клетки в лимфатических узлах и крови. Nat Immunol (2013) 14(2):152–61. дои: 10.1038/ni.2496
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
33. Агата Ю., Кавасаки А., Нисимура Х., Исида Ю., Цубат Т., Ягита Х. и др.Экспрессия антигена PD-1 на поверхности стимулированных Т- и В-лимфоцитов мыши. Int Immunol (1996) 8(5):765–72. дои: 10.1093/интимм/8.5.765
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
34. Кинтер А.Л., Годбаут Э.Дж., МакНалли Дж.П., Серети И., Роби Г.А., О’Ши М.А. и соавт. Общие цитокины гамма-цепи IL-2, IL-7, IL-15 и IL-21 индуцируют экспрессию запрограммированной смерти-1 и ее лигандов. J Immunol (2008) 181(10):6738–46. doi:10.4049/иммунол.181.10.6738
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
35. Пенья-Крус В., Макдонаф С.М., Диас-Грифферо Ф., Крам С.П., Карраско Р.Д., Фриман Г.Дж. PD-1 на незрелых и лиганды PD-1 на мигрирующих клетках Лангерганса человека регулируют активность антигенпрезентирующих клеток. J Invest Dermatol (2010) 130(9):2222–30. doi:10.1038/jid.2010.127
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
36. Thibult ML, Mamessier E, Gertner-Dardenne J, Pastor S, Just-Landi S, Xerri L, et al.PD-1 представляет собой новый регулятор активации В-клеток человека. Int Immunol (2013) 25(2):129–37. дои: 10.1093/интимм/dxs098
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
37. Лим Т.С., Чу В., Сиоу Д.Л., Го С., Йонг Д.П., Сун А.Л. Экспрессия PD-1 на дендритных клетках подавляет функцию CD8+ Т-клеток и противоопухолевый иммунитет. Онкоиммунология (2016) 5(3):e1085146. дои: 10.1080/2162402X.2015.1085146
Полнотекстовая перекрестная ссылка | Академия Google
38.Rodrigues CP, Ferreira ACF, Pinho MP, de Moraes CJ, Bergami-Santos PC, et al. Толерогенные дендритные клетки IDO+ индуцируются тучными клетками, экспрессирующими PD-1. Фронт Иммунол (2016) 7:9. doi:10.3389/fimmu.2016.00009
Полнотекстовая перекрестная ссылка | Академия Google
39. Latchman Y, Wood CR, Chernova T, Chaudhary D, Borde M, Chernova I, et al. PD-L2 является вторым лигандом для PD-1 и ингибирует активацию Т-клеток. Nat Immunol (2001) 2(3):261–8. дои: 10.1038/85330
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
40.Фриман Г.Дж., Уэрри Э.Дж., Ахмед Р., Шарп А.Х. Восстановление истощенных ВИЧ-специфических Т-клеток посредством блокады лиганда PD-1-PD-1. J Exp Med (2006) 203(10):2223–7. doi:10.1084/jem.20061800
Полнотекстовая перекрестная ссылка | Академия Google
41. Браун Дж. А., Дорфман Д. М., Ма Ф. Р., Салливан Э. Л., Муньос О., Вуд Ч. Р. и соавт. Блокада лигандов запрограммированной смерти-1 на дендритных клетках усиливает активацию Т-клеток и продукцию цитокинов. J Immunol (2003) 170(3):1257–66. doi:10.4049/иммунол.170.3.1257
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
42. Пацукис Н., Браун Дж., Петкова В., Лю Ф., Ли Л., Буссиотис В.А. Избирательные эффекты PD-1 на пути Akt и Ras регулируют молекулярные компоненты клеточного цикла и ингибируют пролиферацию Т-клеток. Sci Signal (2012) 5 (230): ra46–46. doi:10.1126/scisignal.2002796
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
43. Chemnitz JM, Parry RV, Nichols KE, June CH, Riley JL.SHP-1 и SHP-2 связываются с основанным на тирозине иммунорецепторным переключающим мотивом запрограммированной смерти 1 при первичной стимуляции Т-клеток человека, но только лигирование рецептора предотвращает активацию Т-клеток. J Immunol (2004) 173(2):945–54. doi:10.4049/jиммунол.173.2.945
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
44. Шеппард К.А., Фитц Л.Дж., Ли Дж.М., Бенандер С., Джордж Дж.А., Вутерс Дж. и соавт. PD-1 ингибирует индуцированное Т-клеточным рецептором фосфорилирование сигналосомы ZAP70/CD3ζ и нижестоящую передачу сигналов к PKCθ. FEBS Lett (2004) 574 (1–3): 37–41. doi:10.1016/j.febslet.2004.07.083
Полнотекстовая перекрестная ссылка | Академия Google
45. Hui E, Cheung J, Zhu J, Su X, Taylor MJ, Wallweber HA, et al. Т-клеточный костимулирующий рецептор CD28 является основной мишенью для ингибирования, опосредованного PD-1. Наука (2017) 355(6332):1428–33. doi:10.1126/science.aaf1292
Полнотекстовая перекрестная ссылка | Академия Google
46. Картер Л., Фаузер Л.А., Юссиф Дж., Фитц Л., Дэн Б., Вуд К.Р. Ингибирующий путь PD-1: PD-L влияет как на CD4+, так и на CD8+ Т-клетки и преодолевается IL-2. Eur J Immunol (2002) 32(3):634–43. doi:10.1002/1521-4141(200203)32:3<634:AID-IMMU634>3.0.CO;2-9
Полнотекстовая перекрестная ссылка | Академия Google
47. Нуриева Р., Томас С., Нгуен Т., Мартин-Ороско Н., Ван Ю, Кая М.К. и соавт. Толерантность или функция Т-клеток определяется комбинаторными костимулирующими сигналами. EMBO J (2006) 25(11):2623–33. doi:10.1038/sj.emboj.7601146
Полнотекстовая перекрестная ссылка | Академия Google
48. Фримен Г.Дж., Лонг А.Дж., Иваи Ю., Бурк К., Чернова Т., Нисимура Х. и соавт.Взаимодействие с иммуноингибиторным рецептором Pd-1 новым членом семейства B7 приводит к негативной регуляции активации лимфоцитов. J Exp Med (2000) 192(7):1027–34. doi:10.1084/jem.192.7.1027
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
49. Nishimura H, Minato N, Nakano T, Honjo T. Иммунологические исследования на мышах с дефицитом PD-1: значение PD-1 как отрицательного регулятора ответов B-клеток. Int Immunol (1998) 10(10):1563–72. дои: 10.1093/интимм/10.10.1563
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
50. Салама А.Д., Читнис Т., Имитола Дж., Ансари М.Дж., Акиба Х., Тусима Ф. и соавт. Критическая роль пути запрограммированной смерти-1 (PD-1) в регуляции экспериментального аутоиммунного энцефаломиелита. J Exp Med (2003) 198(1):71–8. doi:10.1084/jem.20022119
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
51. Нильсен С., Хансен Д., Хасби С., Якобсен Б.Б., Лиллеванг С.Т.Ассоциация предполагаемого регуляторного полиморфизма в гене PD-1 с предрасположенностью к диабету 1 типа. Тканевые антигены (2003) 62(6):492–7. doi:10.1046/j.1399-0039.2003.00136.x
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
52. Веласкес-Крус Р., Ороско Л., Эспиноса-Росалес Ф., Карреньо-Манхаррес Р., Солис-Валлехо Э., Лопес-Лара Н.Д. и соавт. Ассоциация полиморфизмов PDCD1 с системной красной волчанкой в детском возрасте. Eur J Hum Genet (2007) 15 (3): 336–41.дои: 10.1038/sj.ejhg.5201767
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
53. Salvi S, Fontana V, Boccardo S, Merlo DF, Margallo E, Laurent S, et al. Оценка экспрессии CTLA-4 и значимости в качестве нового прогностического фактора у пациентов с немелкоклеточным раком легкого. Cancer Immunol Immunother (2012) 61(9):1463–72. doi: 10.1007/s00262-012-1211-y
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
54. Huang PY, Guo SS, Zhang Y, Lu JB, Chen QY, Tang LQ, et al.Сверхэкспрессия опухолевого CTLA-4 предсказывает плохую выживаемость у пациентов с карциномой носоглотки. Oncotarget (2016) 7(11):13060–8. doi:10.18632/oncotarget.7421
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
55. Ahmadzadeh M, Johnson LA, Heemskerk B, Wunderlich JR, Dudley ME, White DE, et al. Опухолевые антиген-специфические CD8 Т-клетки, инфильтрирующие опухоль, экспрессируют высокие уровни PD-1 и функционально нарушены. Кровь (2009) 114(8):1537–44. дои: 10.1182/кровь-2008-12-195792
Полнотекстовая перекрестная ссылка | Академия Google
56. Chapon M, Randriamampita C, Maubec E, Badoual C, Fouquet S, Wang SF, et al. Прогрессирующая активация PD-1 при первичных и метастатических меланомах связана с притуплением передачи сигналов TCR в инфильтрирующих Т-лимфоцитах. J Invest Dermatol (2011) 131(6):1300–7. doi:10.1038/jid.2011.30
Полнотекстовая перекрестная ссылка | Академия Google
57. Сайто Х., Курода Х., Мацунага Т., Осаки Т., Икегучи М.Повышенная экспрессия PD-1 на CD4+ и CD8+ Т-клетках участвует в уклонении от иммунного ответа при раке желудка. J Surg Oncol (2013) 107(5):517–22. дои: 10.1002/jso.23281
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
58. Xiao X, Lao XM, Chen MM, Liu RX, Wei Y, Ouyang FZ, et al. PD-1hi идентифицирует новую популяцию регуляторных В-клеток в гепатоме человека, которая способствует прогрессированию заболевания. Рак Discov (2016) 6(5):546–59. doi:10.1158/2159-8290.CD-15-1408
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
59.Гордон С.Р., Рой Л.М., Бен В.Д., Грегор Х., Бенсон М.Г., Мелисса Н.М. и др. Экспрессия PD-1 ассоциированными с опухолью макрофагами ингибирует фагоцитоз и опухолевый иммунитет. Природа (2017) 545(7655):495–9. дои: 10.1038/природа22396
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
60. Honda Y, Otsuka A, Ono S, Yamamoto Y, Seidel JA, Morita S, et al. Инфильтрация PD-1-положительными клетками в сочетании с экспрессией PD-L1 в месте опухоли является положительным прогностическим фактором кожной ангиосаркомы. Онкоиммунология (2016) 6(1):e1253657. дои: 10.1080/2162402X.2016.1253657
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
61. Хино Р., Кабашима К., Като Ю., Яги Х., Накамура М., Хондзё Т. и др. Экспрессия опухолевыми клетками лиганда 1 запрограммированной гибели клеток-1 является прогностическим фактором злокачественной меланомы. Рак (2010) 116(7):1757–66. doi:10.1002/cncr.24899
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
62. Хаманиши Дж., Мандай М., Икеда Т., Минами М., Кавагути А., Мураяма Т. и соавт.Безопасность и противоопухолевая активность антитела к PD-1, ниволумаба, у пациенток с резистентным к платине раком яичников. J Clin Oncol (2015) 33(34):4015–22. doi: 10.1200/JCO.2015.62.3397
Полнотекстовая перекрестная ссылка | Академия Google
63. Инман Б.А., Себо Т.Дж., Фригола X, Донг Х., Бергстрах Э.Дж., Франк И. Экспрессия PD-L1 (B7-h2) уротелиальной карциномой мочевого пузыря и БЦЖ-индуцированными гранулемами. Рак (2007) 109(8):1499–505. doi:10.1002/cncr.22588
Полнотекстовая перекрестная ссылка | Академия Google
64.Kim JW, Nam KH, Ahn SH, Park DJ, Kim HH, Kim SH и др. Прогностические последствия экспрессии иммуносупрессивного белка в опухолях, а также инфильтрации иммунных клеток в микроокружении опухоли при раке желудка. Рак желудка (2016) 19(1):42–52. doi: 10.1007/s10120-014-0440-5
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
65. Zhang Y, Kang S, Shen J, He J, Jiang L, Wang W, et al. Прогностическое значение экспрессии запрограммированной гибели клеток 1 (PD-1) или лиганда 1 PD-1 (PD-L1) при раке эпителиального происхождения: метаанализ. Медицина (2015) 94(6):e515. doi:10.1097/MD.0000000000000515
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
66. Ribas A, Hamid O, Daud A, Hodi FS, Wolchok JD, Kefford R, et al. Ассоциация пембролизумаба с опухолевым ответом и выживаемостью среди пациентов с запущенной меланомой. JAMA (2016) 315(15):1600–9. дои: 10.1001/jama.2016.4059
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
67. Ларкин Дж., Чиарион-Силени В., Гонсалес Р., Гроб Дж.Дж., Коуи К.Л., Лао К.Д. и соавт.Комбинация ниволумаба и ипилимумаба или монотерапия при нелеченной меланоме. N Engl J Med (2015) 373(1):23–34. дои: 10.1056/NEJMoa1504030
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
68. Motzer RJ, Escudier B, McDermott DF, George S, Hammers HJ, Srinivas S, et al. Ниволумаб в сравнении с эверолимусом при распространенном почечно-клеточном раке. N Engl J Med (2015) 373(19):1803–13. дои: 10.1056/NEJMoa1510665
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
69.Ansell SM, Iwamoto FM, LaCasce A, Mukundan S, Roemer MGM, Chapuy B, et al. Блокада PD-1 ниволумабом при рецидивирующей или рефрактерной лимфоме Ходжкина. N Engl J Med (2015) 372(4):311–9. дои: 10.1056/NEJMoa1411087
Полнотекстовая перекрестная ссылка | Академия Google
70. Ferris RL, Blumenschein G, Fayette J, Guigay J, Colevas AD, Licitra L, et al. Ниволумаб при рецидивирующем плоскоклеточном раке головы и шеи. N Engl J Med (2016) 375 (19): 1856–67. дои: 10.1056/NEJMoa1602252
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
71.Borghaei H, Paz-Ares L, Horn L, Spigel DR, Steins M, Ready NE и др. Ниволумаб по сравнению с доцетакселом при распространенном неплоскоклеточном немелкоклеточном раке легкого. N Engl J Med (2015) 373(17):1627–39. дои: 10.1056/NEJMoa1507643
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
72. Брамер Дж., Рекамп К.Л., Баас П., Крино Л., Эберхардт В.Е., Поддубская Е. и соавт. Ниволумаб по сравнению с доцетакселом при распространенном плоскоклеточном немелкоклеточном раке легкого. N Engl J Med (2015) 373(2):123–35.дои: 10.1056/NEJMoa1504627
Полнотекстовая перекрестная ссылка | Академия Google
73. Robert C, Schachter J, Long GV, Arance A, Grob JJ, Mortier L, et al. Пембролизумаб против ипилимумаба при прогрессирующей меланоме. N Engl J Med (2015) 372(26):2521–32. дои: 10.1056/NEJMoa1503093
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
74. Nghiem PT, Bhatia S, Lipson EJ, Kudchadkar RR, Miller NJ, Annamalai L, et al. Блокада PD-1 пембролизумабом при распространенной карциноме из клеток Меркеля. N Engl J Med (2016) 374(26):2542–52. дои: 10.1056/NEJMoa1603702
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
75. Гарон Э.Б., Ризви Н.А., Хуэй Р., Лейл Н., Балманукян А.С., Эдер Дж.П. и соавт. Пембролизумаб для лечения немелкоклеточного рака легкого. N Engl J Med (2015) 372(21):2018–28. дои: 10.1056/NEJMoa1501824
Полнотекстовая перекрестная ссылка | Академия Google
76. Reck M, Rodríguez-Abreu D, Robinson AG, Hui R, Csőszi T, et al.Пембролизумаб в сравнении с химиотерапией при PD-L1-положительном немелкоклеточном раке легкого. N Engl J Med (2016) 375 (19): 1823–33. дои: 10.1056/NEJMoa1606774
Полнотекстовая перекрестная ссылка | Академия Google
77. Herbst RS, Baas P, Kim DW, Felip E, Pérez-Gracia JL, Han JY, et al. Пембролизумаб по сравнению с доцетакселом при ранее леченном PD-L1-положительном прогрессирующем немелкоклеточном раке легкого (KEYNOTE-010): рандомизированное контролируемое исследование. Ланцет (2016) 387(10027):1540–50. дои: 10.1016/S0140-6736(15)01281-7
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
78. Le DT, Uram JN, Wang H, Bartlett BR, Kemberling H, Eyring AD, et al. Блокада PD-1 в опухолях с дефицитом репарации несоответствия. N Engl J Med (2015) 372(26):2509–20. дои: 10.1056/NEJMoa1500596
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
79. Armand P, Nagler A, Weller EA, Devine SM, Avigan DE, Chen YB, et al. Отключение иммунной толерантности путем блокады запрограммированной смерти-1 пидилизумабом после аутологичной трансплантации гемопоэтических стволовых клеток при диффузной крупноклеточной В-клеточной лимфоме: результаты международного исследования фазы II. J Clin Oncol (2013) 31(33):4199–206. doi: 10.1200/JCO.2012.48.3685
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
80. Westin JR, Chu F, Zhang M, Fayad LE, Kwak LW, Fowler N, et al. Безопасность и активность блокады PD1 пидилизумабом в комбинации с ритуксимабом у пациентов с рецидивом фолликулярной лимфомы: одногрупповое открытое исследование фазы 2. Lancet Oncol (2014) 15(1):69–77. дои: 10.1016/S1470-2045(13)70551-5
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
81.Риттмейер А., Барлези Ф., Уотеркамп Д., Парк К., Чиардиелло Ф., фон Павел Дж. и др. Атезолизумаб в сравнении с доцетакселом у пациентов с ранее леченным немелкоклеточным раком легкого (OAK): открытое, многоцентровое рандомизированное контролируемое исследование фазы 3. Ланцет (2016) 389(10066):255–65. doi: 10.1016/S0140-6736(16)32517-X
Полнотекстовая перекрестная ссылка | Академия Google
82. Балар А.В., Гальский М.Д., Розенберг Дж.Е., Поулз Т., Петрилак Д.П., Белльмунт Дж. и соавт. Атезолизумаб в качестве терапии первой линии у пациентов с местнораспространенной и метастатической уротелиальной карциномой, не подходящих для лечения цисплатином: одногрупповое многоцентровое исследование фазы 2. Ланцет (2017) 389(10064):67–76. дои: 10.1016/S0140-6736(16)32455-2
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
83. Роберт С., Томас Л., Бондаренко И., О’Дей С., Вебер Дж., Гарб С. и соавт. Ипилимумаб плюс дакарбазин при ранее нелеченой метастатической меланоме. N Engl J Med (2011) 364(26):2517–26. дои: 10.1056/NEJMoa1104621
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
84. Ribas A, Kefford R, Marshall MA, Punt CJ, Haanen JB, Marmol M, et al.Рандомизированное клиническое исследование фазы III, сравнивающее тремелимумаб со стандартной химиотерапией у пациентов с запущенной меланомой. J Clin Oncol (2013) 31(5):616–22. doi: 10.1200/JCO.2012.44.6112
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
85. Antonia SJ, López-Martin JA, Bendell J, Ott PA, Taylor M, Eder JP, et al. Ниволумаб отдельно и ниволумаб плюс ипилимумаб при рецидивирующем мелкоклеточном раке легкого (CheckMate 032): многоцентровое открытое исследование фазы 1/2. Lancet Oncol (2016) 17(7):883–95. дои: 10.1016/S1470-2045(16)30098-5
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
86. Ризви Н.А., Хеллманн М.Д., Снайдер А., Квистборг П., Макаров В., Гавел Дж.Дж., и соавт. Мутационный ландшафт определяет чувствительность к блокаде PD-1 при немелкоклеточном раке легкого. Наука (2015) 348(6230):124–8. doi:10.1126/science.aaa1348
Полнотекстовая перекрестная ссылка | Академия Google
87. Snyder A, Makarov V, Merghoub T, Yuan J, Zaretsky JM, Desrichard A, et al.Генетическая основа клинического ответа на блокаду CTLA-4 при меланоме. N Engl J Med (2014) 371(23):2189–99. дои: 10.1056/NEJMoa1406498
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
88. Tumeh PC, Harview CL, Yearley JH, Shintaku IP, Taylor EJ, Robert L, et al. Блокада PD-1 вызывает ответы путем ингибирования адаптивного иммунного сопротивления. Природа (2014) 515(7528):568–71. дои: 10.1038/природа13954
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
89.Peng W, Liu C, Xu C, Lou Y, Chen J, Yang Y и др. Блокада PD-1 усиливает миграцию Т-клеток в опухоли за счет повышения уровня индуцируемых IFN-γ хемокинов. Cancer Res (2012) 72(20):5209–18. DOI: 10.1158/0008-5472.CAN-12-1187
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
90. Anegawa H, Otsuka A, Kaku Y, Nonomura Y, Fujisawa A, Endo Y, et al. Активация гранзима В и мРНК интерферона-γ в пораженных участках, реагирующих на лечение ниволумабом при метастатической меланоме: отчет о клиническом случае. J Eur Acad Dermatol Venereol (2016) 30(12):e231–2. дои: 10.1111/jdv.13567
Полнотекстовая перекрестная ссылка | Академия Google
91. Nonomura Y, Otsuka A, Ohtsuka M, Yamamoto T, Dummer R, Kabashima K. ADAMTSL5 активируется в тканях меланомы у пациентов с вульгарным идиопатическим псориазом, вызванным ниволумабом. J Eur Acad Dermatol Venereol (2017) 31(2):e100–1. дои: 10.1111/jdv.13818
Полнотекстовая перекрестная ссылка | Академия Google
92. Nonomura Y, Otsuka A, Nakashima C, Seidel JA, Kitoh A, Dainichi T, et al.Клетки Th9 периферической крови являются возможным фармакодинамическим биомаркером эффективности лечения ниволумабом у пациентов с метастатической меланомой. Онкоиммунология (2016) 5(12):e1248327. дои: 10.1080/2162402X.2016.1248327
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
93. Appay V, van Lier RA, Sallusto F, Roederer M. Фенотип и функция субпопуляций Т-лимфоцитов человека: консенсус и проблемы. Цитометрия A (2008) 73(11):975–83. doi:10.1002/cyto.a.20643
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
94.Im SJ, Hashimoto M, Gerner MY, Lee J, Kissick HT, Burger MC и др. Определение CD8+ Т-клеток, обеспечивающих пролиферативный всплеск после терапии PD-1. Природа (2016) 537(7620):417–21. дои: 10.1038/nature19330
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
95. Камфорст А.О., Виланд А., Насти Т., Ян С., Чжан Р., Барбер Д.Л. и соавт. Спасение истощенных CD8 Т-клеток с помощью терапии, направленной на PD-1, зависит от CD28. Наука (2017) 355(6332):1423–7. дои: 10.1126/наука.aaf0683
Полнотекстовая перекрестная ссылка | Академия Google
96. Hugo W, Zaretsky JM, Sun L, Song C, Moreno BH, Hu-Lieskovan S, et al. Геномные и транскриптомные особенности ответа на терапию анти-PD-1 при метастатической меланоме. Сотовый (2017) 168(3):542. doi:10.1016/j.cell.2017.01.010
Полнотекстовая перекрестная ссылка | Академия Google
97. Vétizou M, Pitt JM, Daillère R, Lepage P, Waldschmitt N, Flament C, et al. Противораковая иммунотерапия блокадой CTLA-4 зависит от микробиоты кишечника. Наука (2015) 350(6264):1079–84. doi:10.1126/science.aad1329
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
98. Routy B, Le Chatelier E, Derosa L, Duong CPM, Alou MT, Daillère R, et al. Микробиом кишечника влияет на эффективность иммунотерапии эпителиальных опухолей на основе PD-1. Наука (2018) 359(6371):91–7. doi:10.1126/science.aan3706
Полнотекстовая перекрестная ссылка | Академия Google
99. Абдель-Рахман О., Фуад М. Сетевой метаанализ риска почечной токсичности, связанной с иммунитетом, у онкологических больных, получавших ингибиторы контрольных точек иммунитета. Иммунотерапия (2016) 8(5):665–74. doi:10.2217/imt-2015-0020
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
100. Kato Y, Otsuka A, Miyachi Y, Kabashima K. Обострение вульгарного псориаза при применении ниволумаба при меланоме слизистой оболочки полости рта. J Eur Acad Dermatol Venereol (2016) 30(10):e89–91. дои: 10.1111/jdv.13336
Полнотекстовая перекрестная ссылка | Академия Google
101. Chae YK, Chiec L, Mohindra N, Gentzler R, Patel J, Giles F. Случай сахарного диабета 1-го типа, индуцированного пембролизумабом, и обсуждение диабета 1-го типа, индуцированного ингибитором иммунных контрольных точек. Cancer Immunol Immunother (2016) 66(1):25–32. дои: 10.1007/s00262-016-1913-7
Полнотекстовая перекрестная ссылка | Академия Google
102. Zou W, Wolchok JD, Chen L. Блокада пути PD-L1 (B7-h2) и PD-1 для лечения рака: механизмы, биомаркеры ответа и комбинации. Sci Transl Med (2016) 8(328):328rv4. doi:10.1126/scitranslmed.aad7118
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
103. Фриман-Келлер М., Ким Ю., Кронин Х., Ричардс А., Гибни Г., Вебер Дж.С.Ниволумаб при резецированной и нерезектабельной метастатической меланоме: характеристики иммунозависимых нежелательных явлений и связь с исходами. Clin Cancer Res (2016) 22(4):886–94. doi:10.1158/1078-0432.CCR-15-1136
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
104. Diem S, Kasenda B, Spain L, Martin-Liberal J, Marconcini R, Gore M, et al. Лактатдегидрогеназа сыворотки как ранний маркер исхода у пациентов, получавших анти-PD-1 терапию при метастатической меланоме. Br J Рак (2016) 114 (3): 256–61. doi:10.1038/bjc.2015.467
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
106. Калапре Л., Уорбертон Л., Миллуорд М., Зиман М., Грей Э.С. Циркулирующая опухолевая ДНК (CtDNA) в виде жидкой биопсии меланомы. Cancer Lett (2017) 404 (Приложение C): 62–9. doi:10.1016/j.canlet.2017.06.030
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
107. Lee JH, Long GV, Boyd S, Lo S, Menzies AM, Tembe V, et al.Циркулирующая опухолевая ДНК предсказывает реакцию на антитела против PD1 при метастатической меланоме. Энн Онкол (2017) 28(5):1130–6. дои: 10.1093/annonc/mdx026
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
108. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. Безопасность, активность и иммунные корреляты антитела против PD-1 при раке. N Engl J Med (2012) 366 (26): 2443–54. дои: 10.1056/NEJMoa1200690
Полнотекстовая перекрестная ссылка | Академия Google
109.Weide B, Martens A, Hassel JC, Berking C, Postow MA, Bisschop K, et al. Исходные биомаркеры исхода лечения пациентов с меланомой, получавших пембролизумаб. Clin Cancer Res (2016) 22(22):5487–96. doi:10.1158/1078-0432.CCR-16-0127
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
110. Giacomo AM, Calabrò L, Danielli R, Fonsatti E, Bertocci E, Pesce I, et al. Долгосрочная выживаемость и иммунологические параметры у пациентов с метастатической меланомой, ответивших на лечение ипилимумабом в дозе 10 мкг/кг в рамках программы расширенного доступа. Cancer Immunol Immunother (2013) 62(6):1021–8. дои: 10.1007/s00262-013-1418-6
Полнотекстовая перекрестная ссылка | Академия Google
111. Ishida Y, Otsuka A, Tanaka H, Levesque MP, Dummer R, Kabashima K. HLA-A*26 коррелирует с реакцией на ниволумаб у пациентов с японской меланомой. J Invest Dermatol (2017) 137(11):2443–4. doi:10.1016/j.jid.2017.06.023
Полнотекстовая перекрестная ссылка | Академия Google
112. Gao J, Ward JF, Pettaway CA, Shi LZ, Subudhi SK, Vence LM, et al.VISTA представляет собой ингибиторную контрольную точку иммунного ответа, которая повышается после терапии ипилимумабом у пациентов с раком предстательной железы. Nat Med (2017) 23(5):551–5. дои: 10.1038/nm.4308
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
113. Koyama S, Akbay EA, Li YY, Herter-Sprie GS, Buczkowski KA, Richards WG, et al. Адаптивная устойчивость к терапевтической блокаде PD-1 связана с активацией альтернативных иммунных контрольных точек. Nat Commun (2016) 7:10501.дои: 10.1038/ncomms10501
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
114. Arlauckas SP, Garris CS, Kohler RH, Kitaoka M, Cuccarese MF, Yang KS, et al. Визуализация in vivo выявляет связанный с опухолью путь резистентности, опосредованный макрофагами, при терапии анти-PD-1. Sci Transl Med (2017) 9(389):eaal3604. doi:10.1126/scitranslmed.aal3604
Полнотекстовая перекрестная ссылка | Академия Google
115. Лайнс Дж.Л., Семпере Л.Ф., Бротон Т., Ван Л., Ноэль Р.VISTA — это новый регулятор отрицательной контрольной точки широкого спектра действия для иммунотерапии рака. Cancer Immunol Res (2014) 2(6):510–7. doi:10.1158/2326-6066.CIR-14-0072
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
116. Блейк С.Дж., Дугалл В.К., Майлз Дж.Дж., Тенг М.В., Смит М.Дж. Молекулярные пути: нацеливание на CD96 и TIGIT для иммунотерапии рака. Clin Cancer Res (2016) 22(21):5183–8. doi:10.1158/1078-0432.CCR-16-0933
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
117.Глен М.С., Цуёси Ф., Габриэла М.В., Бенджамин Дж.Б., Хелен Л.В., Джейсон С.Р. и др. TIGIT маркирует истощенные Т-клетки, коррелирует с прогрессированием заболевания и служит мишенью для восстановления иммунитета при ВИЧ и ВИО-инфекции. PLoS Pathog (2016) 12(1):e1005349. doi:10.1371/journal.ppat.1005349
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
118. Spranger S, Koblish HK, Horton B, Scherle PA, Newton R, Gajewski TF. Механизм отторжения опухоли дублетами блокады CTLA-4, PD-1/PD-L1 или IDO включает восстановление продукции IL-2 и пролиферацию CD8(+) T-клеток непосредственно в микроокружении опухоли. J Immunother Cancer (2014) 2:3. дои: 10.1186/2051-1426-2-3
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
119. Antonios JP, Soto H, Everson RG, Moughon D, Orpilla JR, Shin NP, et al. Иммуносупрессивные миелоидные клетки, инфильтрирующие опухоль, опосредуют адаптивную иммунную резистентность через механизм PD-1/PD-L1 при глиобластоме. Neuro Oncol (2017) 19(6):796–807. doi:10.1093/neuonc/now287
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
120.Чамото К., Чоудхури П.С., Кумар А., Сономура К., Мацуда Ф., Фагарасан С. и др. Химические вещества для активации митохондрий синергизируют с блокадой поверхностных рецепторов PD-1 для Т-клеточно-зависимой противоопухолевой активности. Proc Natl Acad Sci USA (2017) 114(5):E761–70. doi:10.1073/pnas.1620433114
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
121. Кесада С.А., Пеггс К.С., Карран М.А., Эллисон Дж.П. Блокада CTLA4 и комбинированная иммунотерапия GM-CSF изменяют внутриопухолевый баланс эффекторных и регуляторных Т-клеток. J Clin Invest (2006) 116(7):1935–45. дои: 10.1172/JCI27745
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
122. Sorensen MR, Holst PJ, Steffensen MA, Christensen JP, Thomsen AR. Аденовирусная вакцинация в сочетании со стимуляцией CD40 и блокадой CTLA-4 может привести к полной регрессии опухоли на модели мышиной меланомы. Вакцина (2010) 28(41):6757–64. doi:10.1016/j.vaccine.2010.07.066
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
123.Чесни Дж., Колличио Ф., Андтбака Р.И., Пузанов И., Гласпи Дж., Мильхем М. и др. Промежуточная безопасность и эффективность рандомизированного (1:1) открытого исследования фазы 2 талимогена лагерпарепвека (Т) и ипилимумаба (I) по сравнению с монотерапией I при нерезецированной меланоме стадии IIIB-IV. Энн Онкол (2016) 27 (приложение_6): 379–400. doi: 10.1093/annonc/mdw379.04
Полнотекстовая перекрестная ссылка | Академия Google
124. Рибас А., Даммер Р., Пузанов И., Вандервальде А., Андтбака Р.И., Мичиелин О. и соавт. Онколитическая виротерапия способствует внутриопухолевой инфильтрации Т-клеток и улучшает иммунотерапию против PD-1. Cell (2017) 170(6):1109.e–19.e. doi:10.1016/j.cell.2017.08.027
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
125. Sivan A, Corrales L, Hubert N, Williams JB, Aquino-Michaels K, Earley ZM, et al. Комменсал Bifidobacterium способствует противоопухолевому иммунитету и повышает эффективность анти-PD-L1. Наука (2015) 350(6264):1084–9. doi:10.1126/science.aac4255
Полнотекстовая перекрестная ссылка | Академия Google
126. Гопалакришнан В., Спенсер К.Н., Нези Л., Рубен А., Эндрюс М.С., Карпинец Т.В., и соавт.Микробиом кишечника модулирует ответ на иммунотерапию анти-PD-1 у пациентов с меланомой. Наука (2018) 359(6371):97–103. doi:10.1126/science.aan4236
Резюме PubMed | Полный текст перекрестной ссылки | Академия Google
MOTUL ATF VI — Motul
Производительность OEM
AUDI G 052 533, VW G 052 533, BMW ATF6, CITROËN AW-1, CITROËN AW-2, DS (CAPSA) AW-2, PEUGEOT AW-2, CITROËN 8 и 9 Speed ATF, DS (CAPSA) 8 & 9 скорость ATF, Peugeot 8 & 9 скорость ATF, CITROëN 68818925AA / AB, DS (CAPSA) 68818925AA / AB, PEUGEOT 68818925AA / AB, CITROëN 68043742AA, DS (CAPSA) 68043742AA, PEUGEOT 68043742AA, CITROëN 05127382, DS (CAPSA) 05127382, PEUGEOT 05127382, DS (CAPSA) AW-1, PEUGEOT AW-1, GENERAL MOTORS 1
39, HONDA 08200-9008, HONDA 08200-9016-A, ACURA 08200-9016A, ZF ZF ZF Lifeguard 9, NISSAN 999MP-MPT-10P, MERCEDES BENZ P/N 00298, JASO 1A, MITSUBISHI MZ320775, GENERAL MOTORS Dex HP, CHRYSLER DEXRON VI Automatic Transmission Fluid, CHRYSLER MS -A5, DAIMLER DEXRON VI Automatic Transmission Fluid, MOPAR PARTS 68218925AA, PETRONAS LUBRICANTS Tutela Transmission AS WUSI8, ISU Transmission AS WUSI8, ISUZU Besco ATF III, FORD MERCON LV, FORD XT-6-QSP (SP), FORD WSS-M2C922-A1, FIAT 9.55550-AV5 для АКПП ZF с 8 и 9 ступенями, NISSAN MATIC P, SUZUKI MATIC S, MITSUBISHI Diamond Queen ATF-PA, MITSUBISHI 4030401, MITSUBISHI ATF-J3, HYUNDAI SP4-M1, KIA SP4-M1, HYUNDAI SPH-IV , KIA SPH-IV, HYUNDAI WS, KIA WS, HYUNDAI NWS-9638, KIA NWS-9638, NISSAN MATIC W, BMW 83 22 2 305 396 (ATF2), BMW 83 22 2 305 397, BMW 83 22 2 413 477 ( ATF7), MINI BMW 83 22 2 413 477 (ATF7), BRILLIANCE ATF PTF-1, PSA Groupe AW-1, AISIN AW-1, AISIN ATF-0WS (кроме вариаторов), AISIN AW-2, SAAB 93 165 147, ХОНДА Хонда Тип 3.0, VOLVO 31 256 774, VOLVO 31 256 775, GENERAL MOTORS AW-1, ZF ZF ZF AA01 500 001, TOYOTA ATF FZ, SUBARU ATF-WS, MERCEDES-BENZ MB 236,12, GENERAL MOTORS GM DEXRON ULV ATF, DAIMLER MS 9602 , BMW 83 22 2 355 599, BMW 83 22 2 355 601, BMW 83 22 2 163 514, BMW 83 22 2 289 720, JASO M315, JALOS Class 1A-LV, ISUZU SCS, MERCEDES-BENZ MB 236.16 (кроме преимуществ FE ), MERCEDES-BENZ MB 236.17 (кроме преимуществ FE), AUDI G 053 001, AUDI G 053 001 A2, AUDI G 060 540 A2, VW G 060 540 A2, VW G 053 001, GENERAL MOTORS GMW16974, GENERAL MOTORS WS, GENERAL MOTORS 88863400, GENERAL MOTORS 88863401, MAZDA MERCON LV, HONDA 08266-99907, HONDA 08266-9904, CHRYSLER DEXRON®-VI Жидкость для автоматических трансмиссий, ACURA ATF Z1 (кроме вариаторов), HONDA ATF Z1 (кроме вариаторов), ACURA ATF DW- 1 (кроме вариаторов), HONDA ATF DW-1 (кроме вариаторов), ACURA 08200-9008A, ACURA 08200-9009A, ACURA 08266-99964, AUDI G 055 005, VW G 055 005, AUDI G 055 162, VW G 055 162 , АУДИ г 060 162, Фольксваген г 060 162, АУДИ г 055 540, Фольксваген г 055 540, БЕНТЛИ ПИ112995ПА , BMW ATF 3+, BMW L12108, BMW 83 22 0 142 516, BMW 83 22 2 152 426, BMW 83 22 9 400 275, BMW 83 22 9 407 738, BMW 83 22 0 397 114, BMW 83 22 0 403 248 , BMW 83 22 0 403 249, BMW 83 22 0 432 807, BMW 83 22 9 407 858, BMW 83 22 9 407 859, CHRYSLER ATF+3, CHRYSLER ATF+4, DAIMLER ATF+4, DAIMLER ATF+3, CHRYSLER МС 9602, Chrysler 68157995AA, Даймлер 68157995AA, CHRYSLER 68157995AB, Даймлер 68157995AB, CHRYSLER 68218057CA, Даймлер 68218057CA, CHRYSLER 68218058CA, Даймлер 68218058CA, CHRYSLER 68218054CA, Даймлер 68218054CA, Chrysler 68218925AA, Даймлер 68218925AA, CHRYSLER ИП И.В. Автоматическая трансмиссия Жидкость, Даймлер ИП И.В. Автоматические Трансмиссионная жидкость, Chrysler 68171866AA, Daimler 68171866AA, Daimler 68171869AA, Chryler 68171869AA, Chryler 68171869AA, Daimler Dexron®-VI Автоматическая коробка передач, Chrysler 68043742AB, Daimler 68043742AB, Ford Mercon Ulv, Ford XT-10-QLVC, General Motors GM Dexron VI, General Motors GM Dexron VI DEXRON III H, GENERAL MOTORS GM DEXRON II D, GENERAL MOTORS 9986153, GENERAL MOTORS 9986333, GEN Eral Motors 88861003, General Motors 88861003, General Motors GM Dexron HP ATF, General Motors GMW16444, General Motors GM 14, General Motors GM
414, Honda 082000-9001, Honda 08200-9005, Honda 08200-9017, Honda ATF-11, Honda B2618-OLE-025, HONDA ATF-TYPE 3.1, КИА СП-ИВ, ХЮНДАИ СП-ИВ, ХЮНДАИ СП-ИВ м, КИА СП-ИВ м, ХЮНДАИ СП-ИВ-РР, КИА СП-ИВ-РР, ХИУНДАИ 040000C90SG, ХЮНДАИ 00232-1904, ХЮНДАИ 04500-00115 , INFINITI MATIC S, NISSAN MATIC S, INFINITI MATIC K, NISSAN MATIC K, NISSAN MATIC J, INFINITI MATIC J, NISSAN MATIC D, INFINITI MATIC D, JAGUAR Jaguar Fluid 8432, JAGUAR 02JDE 26444, LAND ROVER TYK500050, LAND ROVER8 9LR02322 LAND ROVER LR023288, LEXUS ATF WS, TOYOTA ATF WS, LEXUS ATF Type T-IV, TOYOTA ATF Type T-IV, LEXUS JWS 3324, TOYOTA JWS 3324, TOYOTA NWS 9638, LEXUS NWS 9638, LEXUS 00289-ATFWS, TOYOTA 00289- ATFWS, TOYOTA 08886-02303, LEXUS 08886-02303, LEXUS 08886-02305, TOYOTA 08886-02305, LEXUS 08886-80803, TOYOTA 08886-80803, MASERATI 231603, MASERATI M-1375.4, MAZDA ATF FZ, MAZDA 0000-77-122E-01, MERCEDES-BENZ MB 236.14, MERCEDES-BENZ MB 236.15 (кроме преимуществ FE), MERCEDES-BENZ MB 236.41, MERCEDES-BENZ A 001 989 45 03, MERCEDES-BENZ A 001 989 68 03, MERCEDES-BENZ A 001 989 77 03, MERCEDES-BENZ A 001 989 78 03, MITSUBISHI Diamond ATF SP II, MITSUBISHI SP III, MITSUBISHI ATF-MA1, MITSUBISHI ATF SP-IV, MITSUBISHI ATF SPH-IV , MITSUBISHI MZ320762, NISSAN KLE24-00002, NISSAN 999MP-MTS00P, PORSCHE 000 043 304 00, PSA Groupe RP 9734 R7 для трансмиссий EAT6, PEUGEOT RP 9734 R7 для трансмиссий EAT6, CITROËN RP 9734 R7 для трансмиссий EAT6, DS 9734 R7 для трансмиссий EAT6, спецификации материалов SSANGYONG DSIH 5M-66, версия 3, ALLISON TRANSMISSION C-4, ZF ZF ZF M-1375.4, SHELL M-1375.4, ZF ZF ZF M-1375.5, SHELL M-1375.5, SHELL M-1375.6, ZF ZF ZF M-1375.6, ZF ZF ZF TE-ML 11, AML 4G4319A509/AA/S, FUCHS ATF 3292, FUCHS ATF 4400, FUCHS ATF 6000 SL, SHELL L-12108, ZF ZF ZF Lifeguard 6, ZF ZF ZF Lifeguard 6 Plus, ZF ZF ZF Lifeguard 8, ZF ZF ZF S671 090 255, ZF ZF ZF S671 090 281, ZF ZF ZF S671 090 312
.