Кардиотокография плода во время беременности, цена КГТ, расшифровка, оценка
Во время беременности будущая мама должна постоянно посещать гинеколога и проходить диагностические исследования для наблюдения за развитием малыша. Одна из процедур – кардиотокография плода, представляет собой регистрацию частоты сердцебиения и оценку тонуса матки.
В основе метода эффект Доплера, который предполагает получение данных в результате двигательной активности, поэтому важно дождаться шевелений при обследовании. Сегодня это незаменимый этап, позволяющий оценить состояние плода и течение беременности в целом.
Как и когда проводится КТГ?
КТГ плода делается с помощью ультразвукового датчика, который закрепляют на животе там, где достигается оптимальная слышимость сердечного ритма. Ультразвуковые сигналы от сердца плода поступают на устройство, которое мгновенно рассчитывает ЧСС.
Процедура проводится в третьем триместре беременности, а точнее – не раньше 32-й недели.
Результаты КТГ
Расшифровка кардиотокографии плода подразумевает оценку базального ритма сердцебиений, вариабельности сердечного ритма, акцелерации и децелерации. Результаты выражаются в баллах от 0 до 10:
- 8-10 баллов – это норма, которая позволяет делать благоприятные прогнозы на исход родов;
- 7 и менее баллов говорят о лёгком стрессе плода и требуют дальнейшего наблюдения специалистов до родов и в послеродовом периоде;
- оценка менее 4 баллов означает, что есть серьёзные проблемы, поэтому требуется немедленное родоразрешение путём стимуляции или кесарева сечения.
КТГ в Ижевске
Кардиотокография плода в Ижевске проводится в Многопрофильном медицинском центре «Медицея» на современном оборудовании.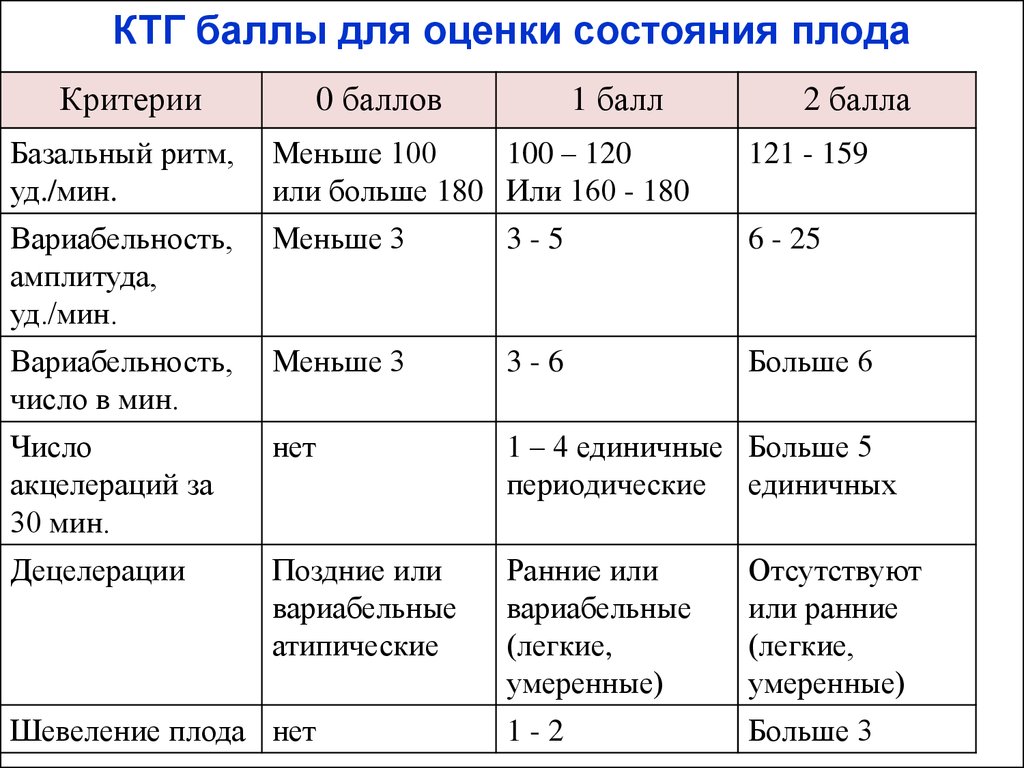 Задать интересующие вопросы и записаться на процедуру можно по телефону +7 (3412) 65-51-51 или заказ обратный звонок на сайте.
Задать интересующие вопросы и записаться на процедуру можно по телефону +7 (3412) 65-51-51 или заказ обратный звонок на сайте.
Пульмонологи назвали показания для КТ легких при подозрении на COVID-19 — РБК
«Для этого должны быть соответствующие показания — например, у человека долго, больше 4–5 дней, держится температура выше 38–39 градусов. И при осмотре, прослушивании врач обнаруживает какие-то изменения в легких, изменения со стороны функции дыхания — тогда он может назначить КТ», — рассказал он.
По словам пульмонолога, профессора кафедры фтизиатрии и пульмонологии лечебного факультета МГМСУ им. А. И. Евдокимова Сергея Бабака, компьютерная томография позволяет «увидеть» структуру легких.
Читайте на РБК Pro
«Показания для КТ — когда мы хотим понять, есть ли структурные изменения в легких, в бронхах, сосудистый компонент, есть ли воспаление в легких, есть ли онкологические проявления. Есть изменения, специфические для вирусного поражения легких, они тоже хорошо видны на КТ», — объяснил врач.
При этом только на основе данных компьютерной томографии пациенту нельзя поставить диагноз, подчеркнул профессор. «Любые методы диагностики бессмысленны без наличия симптомов. Сочетание симптоматики с диагностическими данными повышает точность диагноза. Только с помощью функциональной диагностики поставить диагноз нельзя», — сказал Бабак.
Он отметил, что эту процедуру будет сложно провести пациентам, которые по каким-либо причинам не могут находиться в горизонтальном положении, не могут сделать глубокий вдох, страдают клаустрофобией. Кроме того, компьютерную томографию нельзя проводить беременным и кормящим женщинам.
Ранее главный специалист по лучевой и инструментальной диагностике Минздрава России профессор Игорь Тюрин призвал россиян не делать компьютерную томографию «на всякий случай».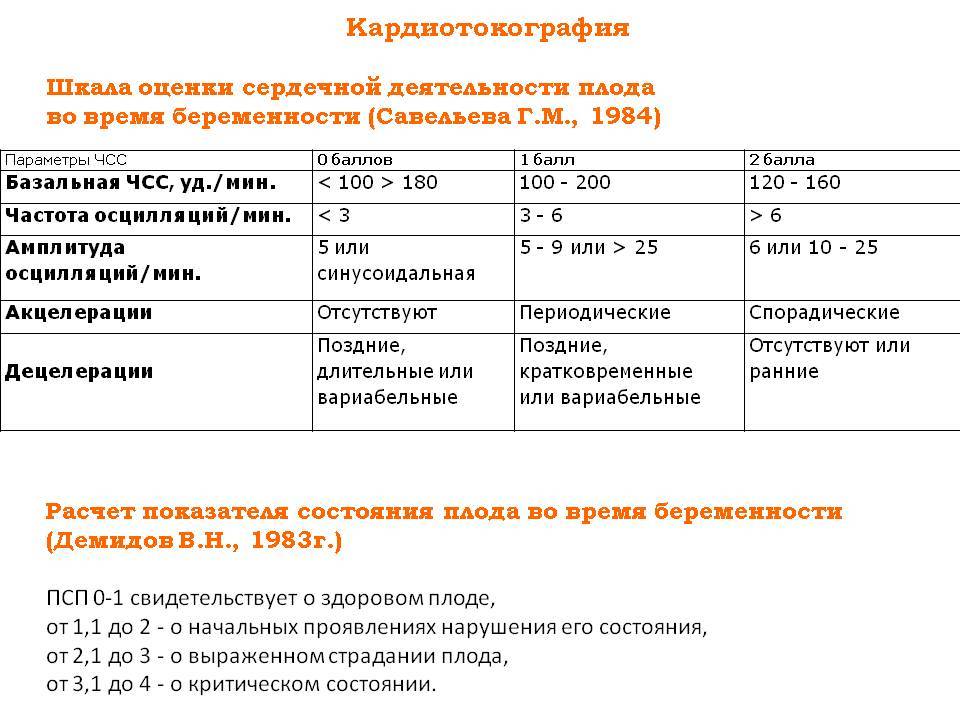 По его словам, эта процедура показана для пациентов с выраженными клиническими симптомами респираторной инфекции (высокая температура, одышка) и пациентов с симптомами ОРВИ, имеющих серьезные факторы риска (выраженный избыток веса, тяжелый диабет, тяжелая сердечная недостаточность).
По его словам, эта процедура показана для пациентов с выраженными клиническими симптомами респираторной инфекции (высокая температура, одышка) и пациентов с симптомами ОРВИ, имеющих серьезные факторы риска (выраженный избыток веса, тяжелый диабет, тяжелая сердечная недостаточность).
Коронавирус
Россия Москва Мир
0 (за сутки)
Заразились
0
0 (за сутки)
Умерли
0 (за сутки)
Заразились
0
0 (за сутки)
Умерли
0 (за сутки)
Заразились
0
0 (за сутки)
Умерли
Источник: JHU, федеральный и региональные оперштабы по борьбе с вирусом
расшифровка, норма, что значит/показывает, как делают
Вынашивание малыша – приятный и очень ответственный период в жизни женщины. Для того чтобы беременность протекала благополучно, роды прошли без осложнений, а малыш появился на свет здоровеньким, за женщиной наблюдают медики на протяжении всего периода гестации.
Когда, как и для чего делают КТГ при беременности
Когда проводят
КТГ проводится всем без исключения женщинам при нормально протекающей беременности трижды в третьем триместре и обязательно во время родов. Проведение кардиотокографии показано с 32 недель. По показаниям исследование возможно провести с 28 недель беремености. Ранее это неинформативно, так как лишь к 28 неделе сердце плода начинает регулироваться вегетативной нервной системой, и его ЧСС начинает реагировать на движения, им совершаемые. Кроме того, к 32 неделе гестации формируется цикличность сна и бодрствования малыша.
Ранее это неинформативно, так как лишь к 28 неделе сердце плода начинает регулироваться вегетативной нервной системой, и его ЧСС начинает реагировать на движения, им совершаемые. Кроме того, к 32 неделе гестации формируется цикличность сна и бодрствования малыша.
Если у женщины имеются проблемы или на предыдущей КТГ были выявлены отклонения, то исследование повторяется чаще.
При внутриутробной гипоксии исследование проводится ежедневно или через день вплоть до нормализации состояния плода или до решения вопроса о необходимости экстренного родоразрешения.
В физиологически протекающих родах КТГ проводится каждые 3 часа. При возникновении осложнений — чаще (период схваток желательно вести под постоянным контролем КТГ).
К показаниям для проведению внеплановой КТГ относятся:
- поздний гестоз (артериальная гипертензия, отеки, белок в моче)
- артериальная гипертензия любой этиологии
- анемия
- многоводие
- маловодие
- многоплодная беременность
- резус-конфликтная беременность
- перенашивание
- угроза преждевременных родов;
- оценка эффективности лечения фетоплацентарной недостаточности и гипоксии плода;
- гипотрофия плода
- тяжелая экстрагенитальная патология матери
При многоплодной беременности исследование проводится отдельно для каждого малыша.
Как делают
Различают 2 способа проведения КТГ: наружный (непрямой) и внутренний (прямой).
Первый способ является самым распространенным. Применяют его без ограничений для всех беременных. У него нет противопоказаний и побочных действий. Во время процедуры датчики помещаются на живот беременной и не доставляют дискомфорта ни ей, ни малышу. Проводят исследование с использованием специального прибора. Он состоит из двух датчиков и устройства записи данных. Оба датчика крепятся на животе беременной специальным ремнем.
Один датчик ультразвуковой. Он позволяет фиксировать частоту сердечных сокращений плода. Второй датчик – тензометрический. Регистрирует сокращения матки. В руку беременной помещается пульт с кнопкой для фиксации шевелений плода.
Оптимальным временем для проведения исследования являются дневные часы с 9 до 14 и вечерние с 19 до 24.
Одно из главных условий для проведения исследования – удобство будущей матери. Ей следует занять комфортное положение сидя на стуле, лежа на спине или боку. На протяжении всей процедуры она не должна испытывать дискомфорта. Длительность исследования составляет обычно 20-40 минут. Это обусловлено периодичностью периодов сна (как правило, не более 30 минут) и бодрствования плода. Регистрацию базального ритма ЧСС плода проводят не менее 20 минут, пока не зафиксируют 2 шевеления длительностью не менее 15 секунд и вызывающие ускорение сердечного ритма на 15 сердечных сокращений в 1 минуту.
Ей следует занять комфортное положение сидя на стуле, лежа на спине или боку. На протяжении всей процедуры она не должна испытывать дискомфорта. Длительность исследования составляет обычно 20-40 минут. Это обусловлено периодичностью периодов сна (как правило, не более 30 минут) и бодрствования плода. Регистрацию базального ритма ЧСС плода проводят не менее 20 минут, пока не зафиксируют 2 шевеления длительностью не менее 15 секунд и вызывающие ускорение сердечного ритма на 15 сердечных сокращений в 1 минуту.
В случае получения неудовлетворительных результатов обычной (нестрессовой) КТГ могут быть назначены специальные тесты (функциональные пробы).
Второй способ используется крайне редко, в основном в момент родов. Для исследования в полость матки вводится катетер или тензодатчик, который фиксирует показатели внутриматочного давления, и ЭКГ-электрод, который крепится к голове плода и регистрирует частоту сердечных сокращений.
Для чего делают КТГ
КТГ проводится для оценки состояния плода в третьем триместре беременности и в процессе (в период схваток и между схватками), выявления угрожающих состояний и решения вопроса об медикаментозных назначения или об экстренном родоразрешении.
В случае получения неудовлетворительных результатов нестрессовой КТГ используются тесты (функциональные пробы), что называется стрессовой КТГ. К данным тестам относятся: окситоциновый, маммарный, акустический, атропиновый и другие.
Расшифровка результата КТГ дает информацию о сердечной деятельности плодаКак подготовиться к процедуре
Результаты КТГ напрямую зависят от состояния матери, поэтому прием пищи перед исследованием должен быть умеренным, в противном случае возросший показатель сахара в крови может привести к чрезмерной активности плода и плохим показателям кардиотокографии. Оптимальным будет результат через два часа после еды.
Искажение результатов исследования может быть следствием:
- потребления большого количества пищи перед обследованием;
- совпадение времени проведения процедуры с периодом сна малыша;
- избыточная масса тела будущей матери;
- чрезмерная активность плода;
- наличие в матке более одного плода;
- неправильное крепление датчиков.

Беременную следует предупредить, что процедура занимает продолжительный период времени и перед ее началом рекомендуется посетить туалет.
Расшифровка результатов
Результат более 9 баллов по 10-балльной шкале – нормаПо итогам исследования врач получает ленту, на которой отображены кривые с разной амплитудой. По ним специалист производит расшифровку результата.
Основные показатели для оценки результата:
- Частота сердечных сокращений (ЧСС), или базальный ритм. В норме в состоянии покоя ЧСС плода находится в границах 120–160 ударов в минуту.
- Высота отклонений от средней частоты сокращений сердечной мышцы. В норме вариабельность не выходит за границы 5–25 ударов в минуту.
- Замедления ЧСС. На ленте кривая уходит вниз, образуя впадину. В норме должны отсутствовать.
- Ускорение ЧСС. На ленте кривая образует зубчатый рисунок. В норме за каждые 10 минут исследования фиксируются две и более акцелерации.
- Сократительная активность матки. Норма составляет не более 15% от ЧСС, длительность от ½ минуты.

Оценка результата производится по 10-балльной шкале Савельевой, где:
- Менее 5 баллов – плохое КТГ. Говорит о наличии острого кислородного голодания – гипоксии. Состояние требует неотложной помощи в виде стимуляции родовой деятельности.
- Показатель 6–7 баллов свидетельствует о начальной стадии кислородного голодания плода. В этом случае рекомендуется лечение в условиях дородового отделения стационара. Через короткий промежуток времени процедура назначается.
- От 8 баллов – норма.
При плохом КТГ важно исключить погрешность измерения, которая может возникнуть в результате неудобной позы беременной во время процедуры.
Одних результатов КТГ недостаточно для постановки диагноза и тем более принятия решения об оперативном родоразрешении. Кроме КТГ существует ряд других исследований, которые могут подтвердить или опровергнуть полученные результаты, например допплер или УЗИ.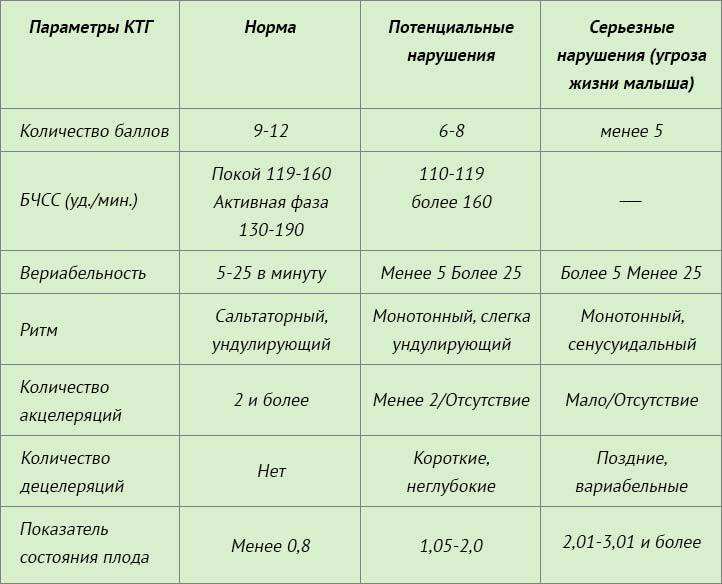
Значимость процедуры
Исследование с помощью кардиотокографа представляет высокую значимость в вопросе оценки состояния плода. Наряду с такими процедурами, как УЗИ, допплерометрия, углубленная электрокардиография, оно позволяет вовремя заподозрить отклонения в сердечно-сосудистой деятельности малыша и принять меры по их коррекции.
При многоплодной беременности, когда оценить работу сердца каждого малыша с помощью стетоскопа не представляется возможным, КТГ – единственный верный способ оценки их состояния.
Если женщина вынашивает однояйцевых близнецов, использование стетоскопа для оценки работы сердца недопустимо, так как полученные результаты будут ложными.
Роды тоже не обходятся без КТГ. С его помощью врач оценивает состояние плода во время схваток и между ними, определяет наиболее подходящий для стимуляции родовой деятельности период, если в этом есть необходимость, рассчитывает дозы препаратов.
Можно ли отказаться от КТГ-исследования при беременности
Некоторые будущие мамы с недоверием относятся к такого рода процедурам. Особенно чувствительным беременным не нравится лежать в одном положении длительное время, других смущают провода.
Нельзя запретить женщине отказаться от процедуры, но только с помощью КТГ можно реально оценить состояние ребенка, записать и учесть его двигательную активность, зафиксировать возможный тонус матки или кислородное голодание.
Для активных будущих мам, которым сложно проводить много времени без движения, современные клиники предлагают беспроводные датчики КТГ и даже датчики, позволяющие производить запись в воде.
Ранняя диагностика возможных патологий позволяет еще на этапе беременности подкорректировать здоровье малыша и благополучно завершить беременность.
Вредит ли кардиотокографическое исследование плоду
В случаях, когда необходим ежедневный мониторинг результатов КТГ, будущие мамы могут волноваться по поводу негативного влияния прибора на ребенка.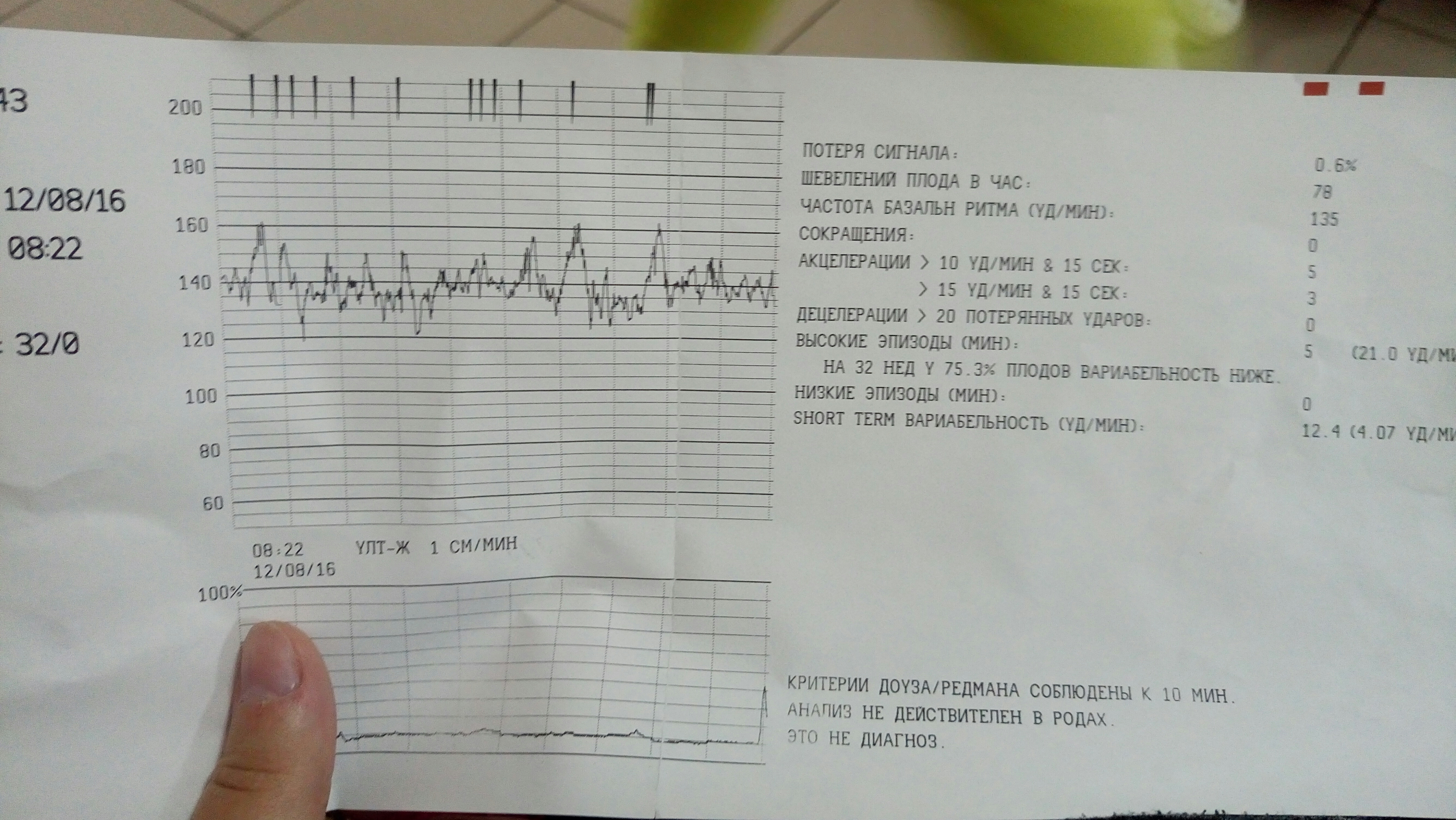 Специалисты уверяют, что аппарат полностью безвреден. Даже ежедневное проведение процедуры не наносит вреда малышу и не доставляет ему дискомфорта.
Специалисты уверяют, что аппарат полностью безвреден. Даже ежедневное проведение процедуры не наносит вреда малышу и не доставляет ему дискомфорта.
Польза внутриутробного обследования плода многократно превышает все возможные риски и опасения будущих матерей по поводу процедуры КТГ. Незначительный дискомфорт у женщины во время процедуры может вызвать лишь длительное отсутствие движения.
Кардиотокография позволяет выявить опасные состояния на самых ранних стадиях, предупредить возможные негативные последствия для плода и беременности в целом и снизить риск их повторного появления. Но следует помнить, что одного исследования мало для постановки точного диагноза. Дополнительно всегда назначаются анализы, УЗИ и допплерометрия.
Видео
Смотрите далее: сердцебиение плода при беременности
Чем МРТ отличается от КТ и какая процедура лучше?
На сегодняшний день магнитно-резонансная томография и компьютерная томография ― наиболее информативные и высокоточные методы инструментальной диагностики. Они создают послойное трехмерное изображение внутренних органов и позволяют делать достоверные заключения о процессах, протекающих в организме пациента.
Они создают послойное трехмерное изображение внутренних органов и позволяют делать достоверные заключения о процессах, протекающих в организме пациента.
Мельчайшие нарушения в тканях будут отражены на снимке и позволят быстро поставить точный диагноз. При этом МРТ и КТ принципиально различаются как в случаях применениях, так и в методах сканирования.
Основные отличия КТ от МРТ
Обе методики визуализируют патологии организма, но если МРТ дает информацию о состоянии мягких тканей, то КТ в большей степени применяется для оценки здоровья костей и других твердых тканей. Ключевые различия МРТ и КТ очевидны, когда понятны принцип действия аппаратов, природа излучения и показания к назначению.
Принцип действия: метод сканирования
Основная разница между исследованиями заключается в методе сканирования:
- В компьютерном томографе применяются рентгеновские лучи, которые проходят через мягкие ткани и отображают плотные структуры. Таким образом, создаются высокоинформативные точные трехмерные послойные изображения. Лучевая нагрузка при проведении компьютерной томографии существенно ниже, чем при выполнении рентгеновского снимка.
- При магнитно-резонансной томографии также создается точное трехмерное послойное изображение органов и тканей, но за счет резонанса атомов водорода в организме человека на магнитное поле, создаваемое томографом.
 Таким образом, создаются высокоинформативные точные трехмерные послойные изображения. Лучевая нагрузка при проведении компьютерной томографии существенно ниже, чем при выполнении рентгеновского снимка.
Таким образом, создаются высокоинформативные точные трехмерные послойные изображения. Лучевая нагрузка при проведении компьютерной томографии существенно ниже, чем при выполнении рентгеновского снимка.Показания к назначению МРТ и КТ
Магнитно-резонансная томография применяется для обследования сосудов, мягких тканей, внутренних органов, головного мозга, нервной системы и лимфатических узлов.
Компьютерная томография помогает обнаружить структурные изменения тканей. В таблице мы приводим список некоторых заболеваний, при которых назначаются эти методы диагностики.
| Показания для назначения МРТ | Показания для назначения КТ |
|---|---|
|
|
 д.
д.Противопоказания к применению МРТ и КТ
Существует ряд противопоказаний к проведению диагностических обследований методами МРТ и КТ. Хотя доза облучения при КТ ничтожно мала, обследование проводят не чаще одного раза в шесть месяцев. Из этого правила могут быть исключения, при жизненной необходимости период между обследованиями может быть сокращен по решению врача.
Из этого правила могут быть исключения, при жизненной необходимости период между обследованиями может быть сокращен по решению врача.
Беременные и кормящие женщины не подлежат диагностике методом КТ из-за негативного воздействия облучения на плод. Пациентам с психическими отклонениями и клаустрофобией также не рекомендуется этот вид обследования из-за особенностей процесса проведения диагностики.
| Противопоказания к МРТ | Противопоказания к КТ |
|---|---|
| Беременность | Беременность |
| Кардиостимулятор | Период грудного вскармливания |
| Слуховой аппарат | Декомпенсированный сахарный диабет |
| Инсулиновая помпа | Тяжелые болезни сердца |
| Несъемные зубные протезы, коронки, мосты | Почечная недостаточность |
| Металлопротезы и осколки | Миелома и плазмоклеточная дискразия |
| Кава-фильтр |
|
При проведении МРТ и КТ пациент должен лежать и не двигаться. Поэтому дети и люди, по состоянию здоровья неспособные лежать без движения в процессе проведения диагностики, а также боящиеся замкнутых пространств пациенты, проходят обследование под наркозом или седацией.
Поэтому дети и люди, по состоянию здоровья неспособные лежать без движения в процессе проведения диагностики, а также боящиеся замкнутых пространств пациенты, проходят обследование под наркозом или седацией.
Как проходит процедура КТ и МРТ?
Ожидание диагностики МРТ и КТ по назначению врача в государственном учреждении здравоохранения в среднем длится месяц. И только экстренные показания (угроза жизни) являются основанием для выполнения обследования вне очереди. В медицинском центре «Адмиралтейские верфи» при наличии назначения врача пройти диагностику на КТ и МРТ можно в день записи.
Обследование на томографе проходит лежа. Врач помогает пациенту разместиться на столе аппарата и выходит из помещения. Исследование занимает от 15 до 20 минут на КТ и от 10 минут до часа на МРТ. Выполнив серию снимков, пациента отпускают. Через полчаса – час ему выдается протокол обследования.
При проведении диагностики с контрастом используется болюсное введение. В нужный момент автоматический инъектор внутривенно вводит контраст пациенту. Использование контраста позволяет создать более детальную картину. Это требуется, когда обычного обследования недостаточно для постановки или уточнения диагноза.
В нужный момент автоматический инъектор внутривенно вводит контраст пациенту. Использование контраста позволяет создать более детальную картину. Это требуется, когда обычного обследования недостаточно для постановки или уточнения диагноза.
Преимущества обследования в медицинском центре «Адмиралтейские верфи»
Многие пациенты сталкиваются с тем, что на УЗИ при выявлении или подозрении на патологию дальнейшее обследование методами КТ или МРТ откладывается на несколько дней или недель.
Если вам требуется пройти обследование оперативно, обращайтесь в отделение инструментальных методов диагностики медицинского центра «Адмиралтейские верфи». Для ускорения и оптимизации алгоритма обследования пациентов мы объединили отделение функциональной и лучевой диагностики.
Для проведения дополнительных исследований нашим пациентам не нужно перезаписываться на последующие даты. Расписание специалистов клиники построено таким образом, что при необходимости мы можем провести дообследование сразу же.

В каждом конкретном случае, решение о выборе того или иного метода диагностики принимает врач: именно он решает, что лучше – МРТ или КТ. Специалисты медицинского центра «Адмиралтейские верфи» ответят на ваши вопросы и подберут адекватный метод диагностики. Для этого позвоните по номеру телефона, указанному на сайте, или оставьте заявку в форме обратной связи. Давайте заботиться о вашем здоровье вместе!
Кардиотокография в Москве — кардиотокография в частной клинике «ВитаМед».
Кардиотокография плода – важная диагностика во время беременности. Кардиотокография предполагает беспрерывную регистрацию двигательной активности, сердцебиения ребенка, сокращений мышц матки.
Кардиотокография плода проводится для выявления симптомов анемии ребенка, гипоксии, отклонений в работе сердца. Процедура позволяет выявлять проблемы фетоплацентарной недостаточности и маловодия.
Когда и зачем производится КТГ плода при беременности
Обследование КТТ плода назначается, чтобы акушер гинеколог мог своевременно выявлять нарушения в состоянии плода, и рекомендовать необходимые шаги для адекватного лечения. Данное исследование становится основой для оценки эффективности терапии. В случае плохого состояния ребенка, кардиотокография при беременности, допплерометрия и УЗИ позволят установить подходящие сроки и тактику для предстоящих родов. Кардиотокография плода позволит планировать подходящие действия для безопасности беременности и родов.
Данное исследование становится основой для оценки эффективности терапии. В случае плохого состояния ребенка, кардиотокография при беременности, допплерометрия и УЗИ позволят установить подходящие сроки и тактику для предстоящих родов. Кардиотокография плода позволит планировать подходящие действия для безопасности беременности и родов.
Показания для выполнения КТГ с расшифровкой
- КТГ при беременности записывается один раз, если ничего не беспокоит, и врач не считает нужным делать запись.
- При патологическом варианте записи. Если при записи сердцебиения выявлены признаки патологии, нужно будет повторить исследование КТГ при беременности с расшифровкой.
- При неблагоприятном течении прошлых беременностей.
- В случае нарушения в поведении будущего ребенка.
- При болезнях самой матери. К примеру, при случаях ОРВИ, гриппа либо острых состояний, ухудшающих общее самочувствие женщины.
- При перенесенных и вылеченных состояниях будущего ребенка.
- В случае гестоза во время беременности.
- При хронических инфекциях беременной женщины.
- Для женщин, продолжающих курить и употреблять алкоголь во время беременности.
- При хронических болезнях внутренних органов беременной женщины.

Противопоказания к кардиотокографии при беременности
Кардиотокография при беременности не имеет противопоказаний. На практике подтверждена абсолютная безопасность процедуры для каждой женщины и ребенка. Полностью безболезненная процедура. Данный информативный метод диагностики позволяет убедиться в отсутствии угроз и опасностей для плода.
Как подготовиться к кардиотокографии
Кардиотокография плода не требует специальной подготовки. Но для объективности полученных результатов женщине важно принять максимально удобное положение. Как правило, беременной женщине предлагается сесть, откидываясь назад на спинку стула или лечь полубоком.
За 8-12 часов до выполнения процедуры следует отказаться от применения спазмолитиков, успокоительных, обезболивающих и других препаратов, влияющих на результаты диагностики. Во время процедуры женщина должна быть здоровой, поскольку ОРВИ, ОРЗ и прочие инфекционные болезни могут нарушить данные диагностики.
Во время процедуры женщина должна быть здоровой, поскольку ОРВИ, ОРЗ и прочие инфекционные болезни могут нарушить данные диагностики.
Как проводится кардиотокография в ВитаМед
Для кардиотокографии плода используется специальная аппаратура, объединяющая датчик, подключенный к устройствам записи данных. Врач прикладывает к животу стетоскоп для контроля сердцебиения ребенка.
Так во время процедуры кардиотокографии выявляется место лучшего прослушивания сердцебиения плода. Затем кладется на данную область ультразвуковой датчик, который крепится к туловищу ремнем. Данный датчик способен фиксировать сердечную активность ребенка. Также производится пристегивание и второго датчика в животу – однако в области дна матки.
Воздушная прослойка между датчиком и кожей живота может нарушить данные исследования. Чтобы её удалить, используется полностью безопасный гель.
Женщина на пульте может нажимать кнопку каждый раз, когда чувствует шевеления ребенка – для лучшего оценивания изменения частоты сердечных сокращений малыша в период своей активности.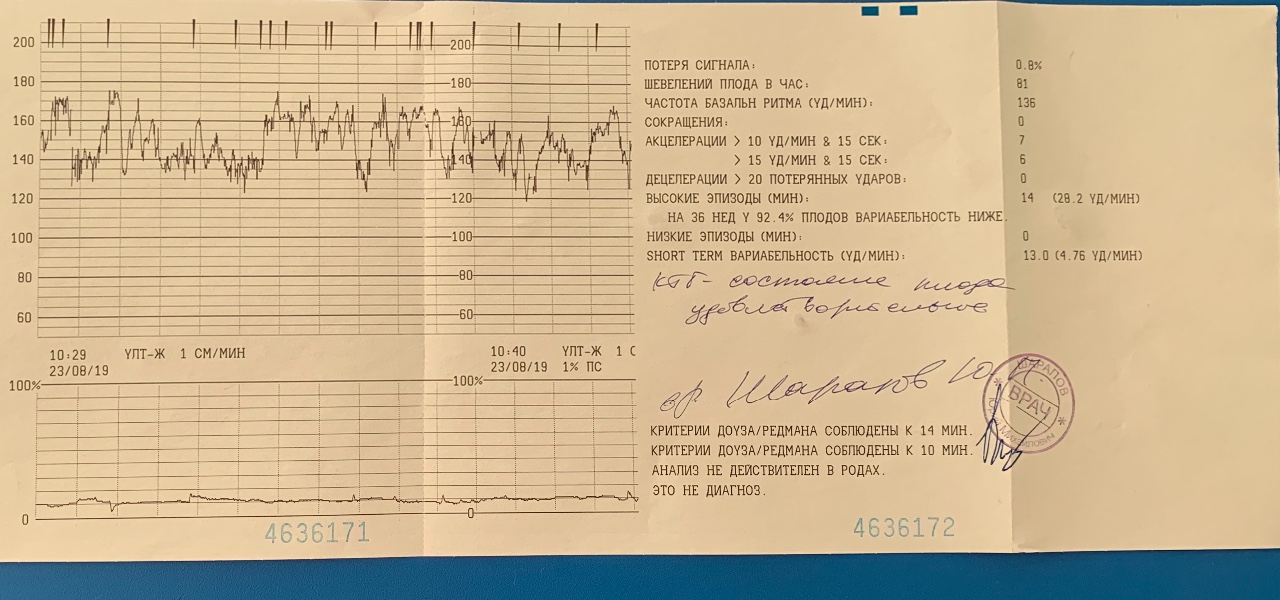
Обычно длительность процедуры кардиотокографии составляет 40, 60 либо 90 минут. Однако конкретная продолжительность процедуры может различаться в зависимости от индивидуальных показаний – часто достаточно меньше времени для выводов о состоянии и самочувствии ребенка.
Расшифровка результатов кардиотокографии при беременности
Кардиотокография с расшифровкой результатов обычно предполагает выдачу результатов на пленке – с 2 графиками, тонусом матки и сердцебиением ребенка. Они обычно сопровождаются описанием КТГ либо с расшифровкой на компьютере.
Верхний график – тахограмма ребенка, с отражением динамики изменений сердцебиения за единицу времени.
Кардиотокограмма оценивается по 5 критериям, каждый из них устанавливается от 0 до 2 баллов – частота, базальный ритм, также амплитуда отклонений от базального ритма, данные децелерации и акцелерации частоты сердцебиений.
9-20
Лечащие врачи:
Первичный приемОнкологАкушер — гинекологМаммологКардиологМассажистПроктологФлебологХирургУЗИ
Что это — КТГ: расшифровка при беременности
Беременность — это чудесный период в жизни женщин. Каждая будущая мама тщательно готовится к встрече со своим малышом. Это заключается в том, что на протяжении всей беременности женщины заботятся не только о своем здоровье, но и о ребенке, который развивается внутри них. Современные технологии позволяют создавать все условия для поддерживания здоровой беременности. Для этого женщины в интересном положении регулярно сдают анализы и проходят обследования. Более подробно остановимся на том, что такое КТГ, как проводят это исследование, для чего оно вообще нужно.
Каждая будущая мама тщательно готовится к встрече со своим малышом. Это заключается в том, что на протяжении всей беременности женщины заботятся не только о своем здоровье, но и о ребенке, который развивается внутри них. Современные технологии позволяют создавать все условия для поддерживания здоровой беременности. Для этого женщины в интересном положении регулярно сдают анализы и проходят обследования. Более подробно остановимся на том, что такое КТГ, как проводят это исследование, для чего оно вообще нужно.
Краткая характеристика процедуры
В медицине аббревиатура КТГ обозначает кардиотокографию, которая является одним из самых ценных и необходимых исследований в течение всей беременности. Поясним, что показывает КТГ. Это процедура, которая позволяет получить сведения о работе сердечка малыша: какова частота биения и сердечная деятельность. Кроме того, при помощи процедуры можно обнаружить двигательную активность плода и работу матки женщины, определить ее сокращения.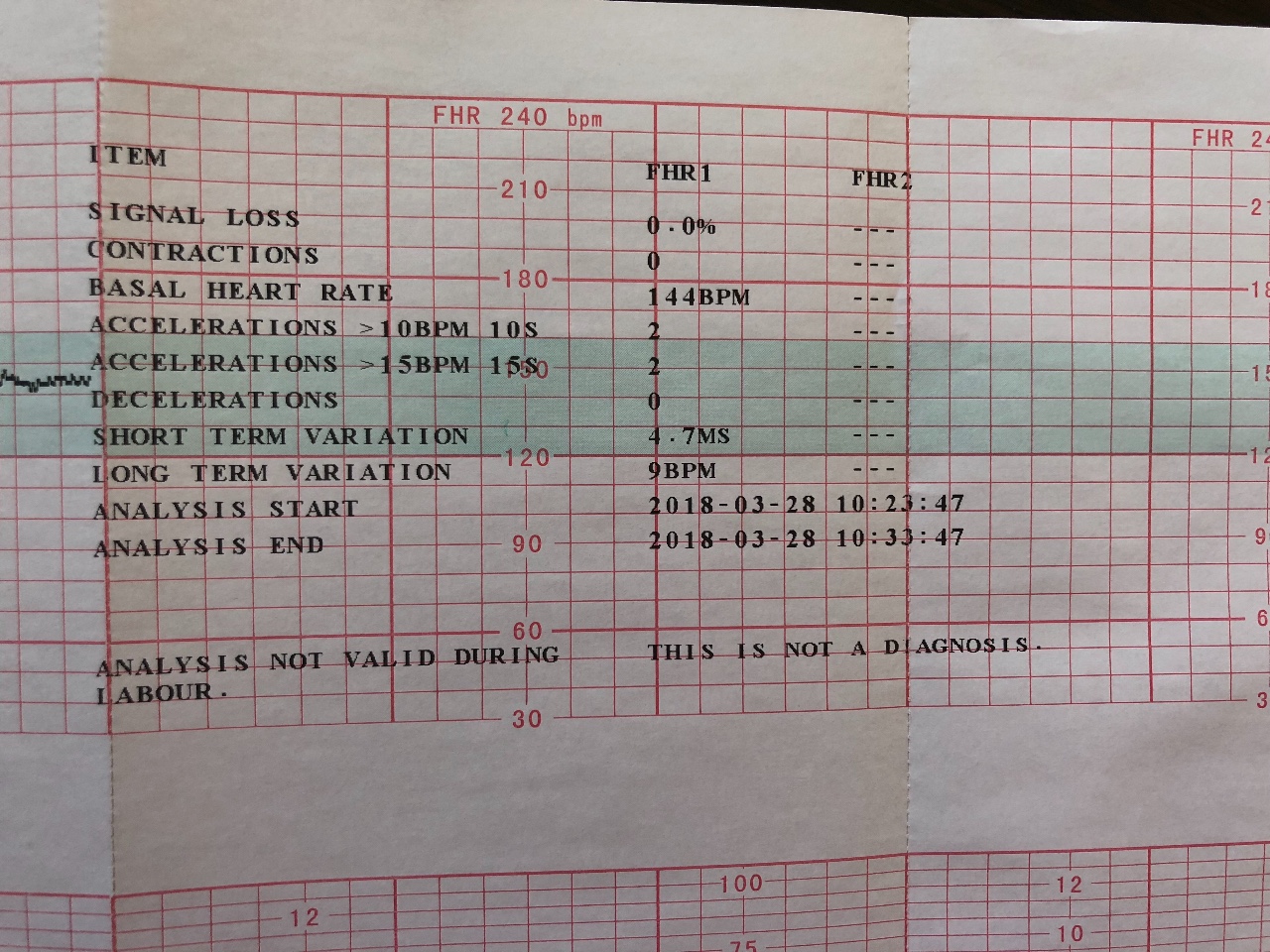 При помощи данного обследования можно также понять реакцию малыша на изменения в организме матери.
При помощи данного обследования можно также понять реакцию малыша на изменения в организме матери.
Что показывает КТГ еще? Этот исследование подтверждает или опровергает опасные явления, которые могут возникнуть во время беременности как для мамы, так и для ребенка. О каких явлениях идет речь? Это гипоксия плода (нехватка кислорода), многоводие, инфекции, которые развиваются внутри матки, преждевременная зрелость плаценты, отклонения в развитии сердца и сосудистой системы плода.
Исходя из всего вышесказанного, мы делаем вывод, что КТГ беременным делать полезно и важно, но обратим внимание, что совсем не обязательно. Исследование покажет и даст оценку не только развития ребенка, но и состояния мамы, а также их совместимости.
Когда стоит делать КТГ
Определившись, что такое КТГ, рассмотрим, на каком сроке этот анализ делают. В качестве начального порога обозначим цифру — 28 неделя беременности. С этого момента можно начинать проводить исследование, но результаты на этом этапе могут быть не совсем точными.
Лучшим сроком является 32 неделя и далее. В этот период беременности у малыша возникают и развиваются нервные импульсы и мышечные сокращения, которые легко отслеживаются аппаратурой. Кроме того, у ребенка формируется цикл, состоящий из фаз отдыха и покоя, поэтому результаты обследования будут более точными и результативными.
КТГ в период родов
По сути, что такое КТГ? Это уникальная процедура, которую можно сделать не только в процессе беременности, но и в момент родов. Алгоритм проведения исследования будет абсолютно такой же, как во время беременности, но есть некоторые особенности:
- Во-первых, если родовая деятельность стабильна, а проблем нет, то все время лежать под прибором роженице не нужно. Можно подключать его время от времени для оценки состояний ребенка и женщины.
- Во-вторых, если используются методы стимуляции родов, то желательно выполнять исследование чаще. Именно так можно определить, помогают ли средства стимуляции, как идет родовой процесс. Получив информацию с помощью КТГ о сердцебиении плода и сокращениях матки, врачи могут рассчитать дозировку препаратов и выстроить правильную тактику.
 Получив информацию с помощью КТГ о сердцебиении плода и сокращениях матки, врачи могут рассчитать дозировку препаратов и выстроить правильную тактику.
Получив информацию с помощью КТГ о сердцебиении плода и сокращениях матки, врачи могут рассчитать дозировку препаратов и выстроить правильную тактику.Если аппаратуры нет и проведение процедуры невозможно в родильном зале, не стоит волноваться. В теории и на практике применяются другие исследования, благодаря которым акушеры могут помочь родить женщине здорового ребенка. КТГ — это лишь одна из многочисленных методик оценки состояния мамы и малыша.
Внутреннее КТГ
Особенным является внутреннее обследование, которое делается не так часто. Что такое КТГ в полости матки? Это узкий вид обследования, который применяется только в процессе родов. Его можно применять только тогда, когда плодный пузырь уже лопнул, то есть его целостность нарушена. Проводится процедура в отдельных случаях, когда есть в этом острая необходимость, например, сомнения в нормальном состоянии плода, подозрение на прекращение сердцебиения или ситуации, когда ребенок перестал реагировать на сокращения матки мамы. Для решения всех этих проблем в полость матки вводится специфический катетер, на конце которого есть электроды. Они прикрепляются к поверхности головы плода. С их помощью определяется работа организма ребенка, динамика многих процессов, выстраивается тактика дальнейших родов, принимаются жизненно важные решения.
Для решения всех этих проблем в полость матки вводится специфический катетер, на конце которого есть электроды. Они прикрепляются к поверхности головы плода. С их помощью определяется работа организма ребенка, динамика многих процессов, выстраивается тактика дальнейших родов, принимаются жизненно важные решения.
Аппарат для КТГ
Ознакомившись с преимуществами обследования, у многих возникнет вопрос, как делают КТГ? С помощью специального аппарата, который состоит из двух датчиков. Они связаны между собой устройством, которое записывает звук. Первый датчик отслеживает сердцебиение ребенка, а второй — сокращения матки и рефлексы ребенка. Если с устройством разобрались, перейдем к тому, как делают КТГ.
Оба датчика прикрепляют к поверхности живота будущей мамы. При этом каждый из них смазывается гелем, который применяется и при УЗИ. В руки беременной дается пульт, на котором есть одна кнопка. Ее нужно нажимать тогда, когда женщина чувствует шевеление ребеночка. В этот момент его активности анализируются изменения сердцебиения плода.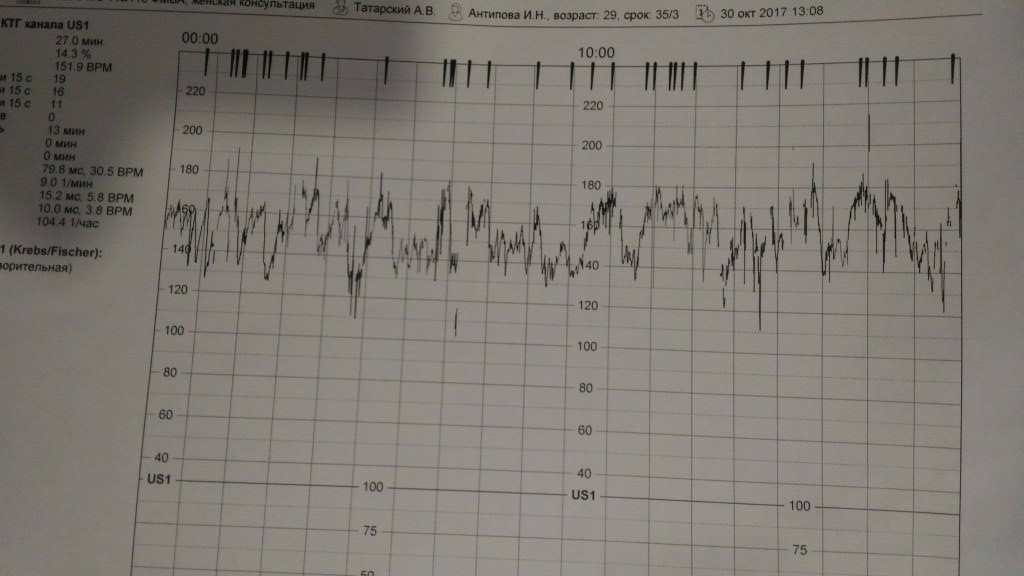 Как и в случае с кардиограммой, все результаты отражены на бумаге в виде кривой. Ее анализирует врач, но современные приборы самостоятельно считают результаты и выставляют баллы.
Как и в случае с кардиограммой, все результаты отражены на бумаге в виде кривой. Ее анализирует врач, но современные приборы самостоятельно считают результаты и выставляют баллы.
Подготовка беременной
Мы определили, что такое КТГ. Вы поняли, что это обследование немного похоже на УЗИ, поэтому особенной подготовки к нему не потребуется. Следует знать, что процедура длится на протяжении примерно 60 минут. Поэтому можно взять с собой книгу или что-то другое, чтобы время прошло быстрее и комфортнее. Можно перекусить перед процедурой, так как нельзя быть голодной. Но и переедать не стоит.
Самое необходимое, что вы должны взять с собой, — хорошее настроение. Желательно перед КТГ настроиться на положительный лад. Важно занять удобное положение. Можно лечь, но не на спину, а на бок. Кроме того, можно проходить процедуру полулежа.
Как быть, если ребенок не хочет проявлять активность, поэтому процесс исследования затягивается? Не стоит беспокоиться на этот счет. Нужно просто что-то съесть. Это должна быть не тяжелая пища, а легкий перекус. В этот момент малыш наверняка начнет двигаться, значит, результаты не заставят себя ждать.
Это должна быть не тяжелая пища, а легкий перекус. В этот момент малыш наверняка начнет двигаться, значит, результаты не заставят себя ждать.
Если вас беспокоит вопрос, где сделать КТГ, не переживайте. Врач, ведущий беременность, обязательно подскажет. Зачастую это делается в женской консультации или в платных центрах.
Результаты. Основные параметры
Как мы говорили ранее, результаты выдаются на современной аппаратуре в виде баллов. Поэтому важен процесс расшифровки КТГ плода. Все показатели имеют свою оценку, которая колеблется от 0 до 2. К таким критериям относятся базальный ритм, амплитуда, процесс учащения сердцебиения или его снижения и другие важные данные. Все баллы по каждому положению складываются. В результате получается сумма. О ней специалист обязательно говорит будущей маме и делает устное пояснение, сравнивая показатели с нормой КТГ плода:
- Результат от 9 до 12 единиц означает, что у малыша все хорошо, он нормально реагирует на сокращения матки, отклонений нет. Дальнейшее наблюдение беременности может протекать в плановом режиме.
- От 6 до 8 баллов означает, что у малыша есть признаки гипоксии в умеренной форме. Получив такие результаты, специалист назначает повторное обследование, которое проводится на следующий день или через день. Если диагноз подтвердится, назначается лечение.
- 5 и меньше баллов указывают на ярко выраженное кислородное голодание плода. Это является показанием для экстренных мероприятий. Иногда это кесарево сечение.
 Дальнейшее наблюдение беременности может протекать в плановом режиме.
Дальнейшее наблюдение беременности может протекать в плановом режиме.Базальный ритм
Прибор также фиксирует частоту сердцебиения ребенка. В данном случае не требуется расшифровки КТГ плода. Если цифра колеблется в районе от 110 до 160 ударов, беспокоиться не стоит, так как это норма. В медицине показатель называется базальным ритмом. На графике ярко иллюстрируются падения и взлеты, поэтому врач оценивает картину, не обращая внимания на числовые показатели. Не беспокойтесь, если результаты для вас непонятны, доверьтесь специалисту. Помните о том, что важно не только то, где сделать КТГ, но и кто его делает. Хороший специалист всегда предоставит пояснение. Если его нет, то обратитесь к гинекологу, ведущему беременность. Он обязательно вам все разъяснит.
Хороший специалист всегда предоставит пояснение. Если его нет, то обратитесь к гинекологу, ведущему беременность. Он обязательно вам все разъяснит.
Опасно ли КТГ
Эта процедура абсолютно безболезненна и безопасна. Она не имеет противопоказаний. КТГ не навредит малышу и не отразится негативно на здоровье мамы. Напротив, результаты помогут ликвидировать угрозу или предупредить появление отклонений и заболеваний. Напомним, что результаты необходимо анализировать в совокупности с другими данными исследований, анализов крови, УЗИ. Не переживайте, если вам выписывают направление на КТГ каждую неделю. Возможно, ваш гинеколог хочет что-то перепроверить. Особенно часто так бывает, если результаты не идеальны. Никакого облучения ребенка и действия вредных импульсов не происходит, поэтому смело проходите эту процедуру.
Кому назначают КТГ
Многих беременных беспокоит вопрос, все ли должны проходить такое обследование. Некоторые спрашивают, почему им его не назначают.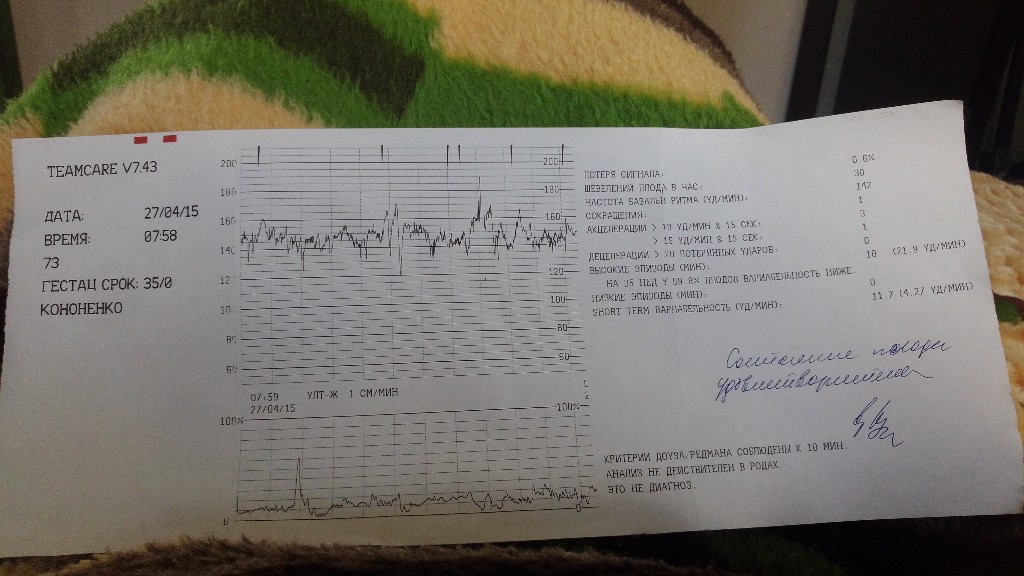 Спешим вас успокоить, КТГ назначается в том случае, если для этого есть показания. Как мы определили ранее, с помощью обследования можно обнаружить патологии, например, гипоксию или многоводие. Если по результатам УЗИ, анализов крови и иных исследований гинеколог, который ведет вашу беременность, отмечает наличие отклонений или угрозы, он направит на КТГ.
Спешим вас успокоить, КТГ назначается в том случае, если для этого есть показания. Как мы определили ранее, с помощью обследования можно обнаружить патологии, например, гипоксию или многоводие. Если по результатам УЗИ, анализов крови и иных исследований гинеколог, который ведет вашу беременность, отмечает наличие отклонений или угрозы, он направит на КТГ.
Если беременность протекает нормально, а все анализы без отклонений, в проведении КТГ нет необходимости. Хотя некоторые врачи перестраховываются, поэтому с 32 недели беременности направляют будущую маму проводить обследование. Главное, не волноваться и не беспокоиться.
Как показывает практика, КТГ могут не делать на протяжении всей беременности, зато в период родов подключают аппарат и отслеживают процесс. В этом нет ничего плохого. Напротив, в некоторых случаях, например, требуется стимуляция, КТГ окажет врачам большую помощь.
Значение КТГ
На протяжении всей беременности женщине придется сделать множество анализов, пройти ряд обследований, многие из которых необходимо будет проводить регулярно. Все это делается лишь для того, чтобы процесс вынашивания ребенка проходил без отклонений, ведь рождение здорового ребеночка — истинное счастье.
Все это делается лишь для того, чтобы процесс вынашивания ребенка проходил без отклонений, ведь рождение здорового ребеночка — истинное счастье.
Кардиотокография — это один из многочисленных видов обследований, которые назначаются беременным. Все чаще КТГ используют для отслеживания динамики развития ребенка в третьем триместре. Ведь неблагоприятная окружающая среда, стрессы, неправильный образ жизни ставят под угрозу полноценное развитие малыша. Не нужно забывать и о том, что многие беременные продолжают работать, что также отражается на состоянии их организма. В данной статье мы не только определили понятие исследования, но и описали расшифровку результатов, отметили, на каком сроке КТГ можно начинать, когда проводить его более результативно. Также нами были определены нюансы и особенности процедуры.
Ктг как должно выглядеть хорошее. Расшифровка ктг при беременности
Существуют некоторые методы ультразвуковой диагностики состояния плода у будущих мам, которые считаются абсолютно безвредными для малыша. Одним из таких методов является кардиотокография (КТГ) при беременности, норма или отклонение от нее по отдельным показателям этого метода диагностики может выявляться по различным шкалам. На основе полученных результатов можно рассматривать вопрос состояния плода и корректировать при необходимости ведение беременности.
Одним из таких методов является кардиотокография (КТГ) при беременности, норма или отклонение от нее по отдельным показателям этого метода диагностики может выявляться по различным шкалам. На основе полученных результатов можно рассматривать вопрос состояния плода и корректировать при необходимости ведение беременности.
КТГ не относится к обязательным процедурам, поэтому врач может вообще не направлять будущую маму на это исследование, однако если возникают какие-то опасения, несколько раз процедуру обязательно проводят.
Особенности процедуры КТГ
Такой способ диагностики используют в третьем триместре беременности, обычно с 32 недели. Некоторые врачи назначают это исследование на 28 неделе, но критерии для диагностики считаются достоверными именно с 32 недели, поскольку к этому времени у плода стабилизируется цикл активность-покой.
С помощью КТГ плода оценивается состояние будущего ребенка во время естественной двигательной активности. Если малыш никак себя не проявляет или вообще спит, скорее всего, процедуру перенесут или проведут еще раз – в этом нет ничего угрожающего.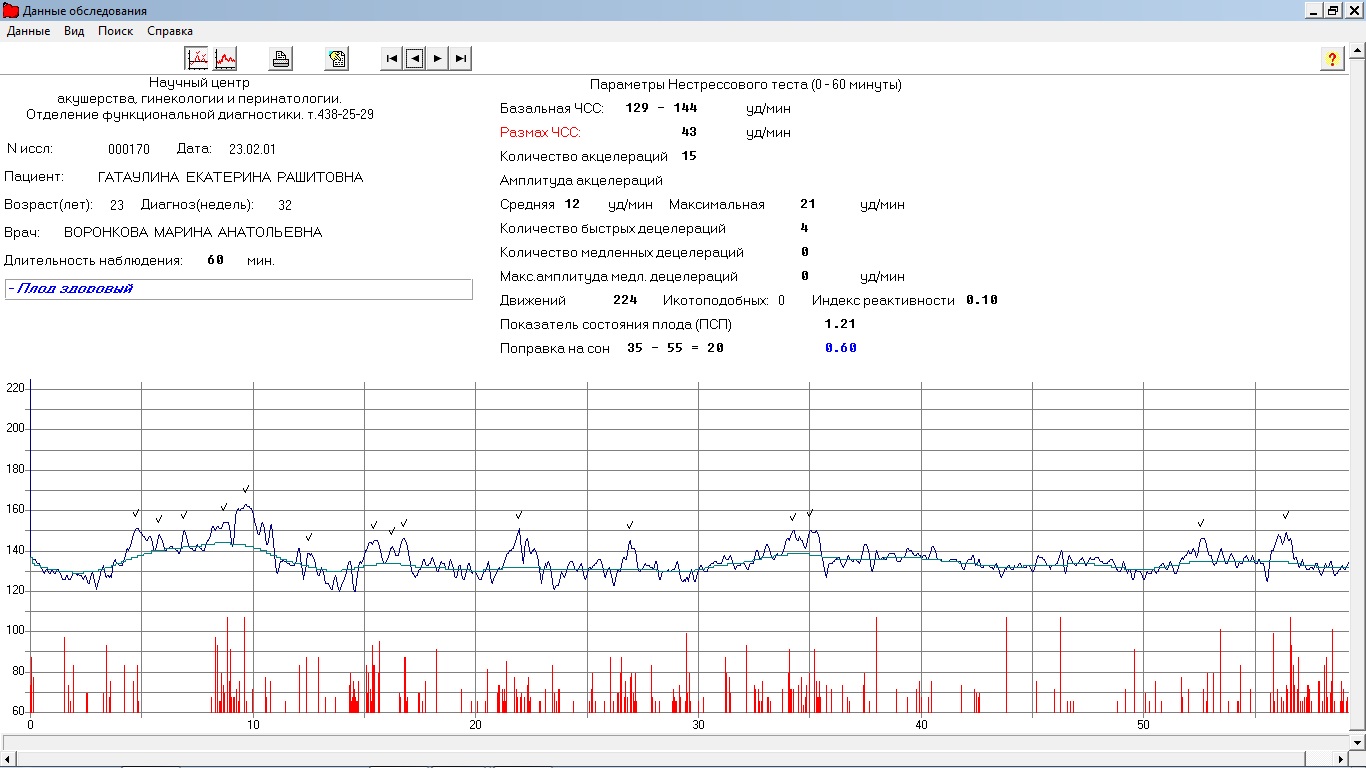 Так как метод абсолютно безвредный для ребенка, можно проводить его бесконечное количество раз.
Так как метод абсолютно безвредный для ребенка, можно проводить его бесконечное количество раз.
Если рассматривать вопрос, в каких случаях или когда делают КТГ при беременности , то обычно это происходит в следующих состояниях:
- выявленная на УЗИ патология со стороны плаценты,
- подозрение на задержку развития плода,
- возможность ,
- уменьшение активности плода,
- сопутствующие хронические заболевания у беременной,
- или ,
- отклонения в прошлой КТГ,
- обвитие плода , выявленное на УЗИ.
Никакой специальной подготовки перед процедурой не существует. Однако каждая женщина хочет быть готовой к тому, что ее ожидает. Если назначили КТГ при беременности, как подготовиться к ней, каждой подскажет здравый смысл. Так как исследование проводится около 40-60 минут, стоит настроиться на долгое времяпрепровождение: взять легкий перекус (яблоко, хлеб, шоколад), плед и подушечку для удобства. Обязательно перед процедурой нужно сходить в туалет, иначе терпеть придется долго, а результаты окажутся недостоверными.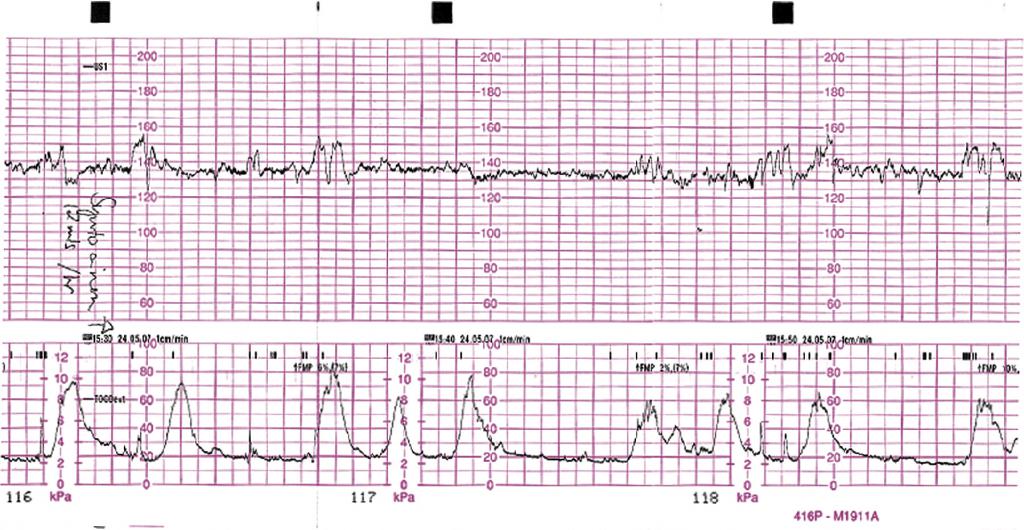
Будущую маму уложат на кушетку или в удобное кресло (нужно находиться в состоянии полулежа или лежа на боку, на спине лежать нельзя), прикрепят датчик на живот, информация от которого пойдет в электронный блок. Врач получает и изучает все данные, которые отражаются на кривой. После исследования врач пишет заключение, которое отдается беременной.
Результаты КТГ плода: расшифровка
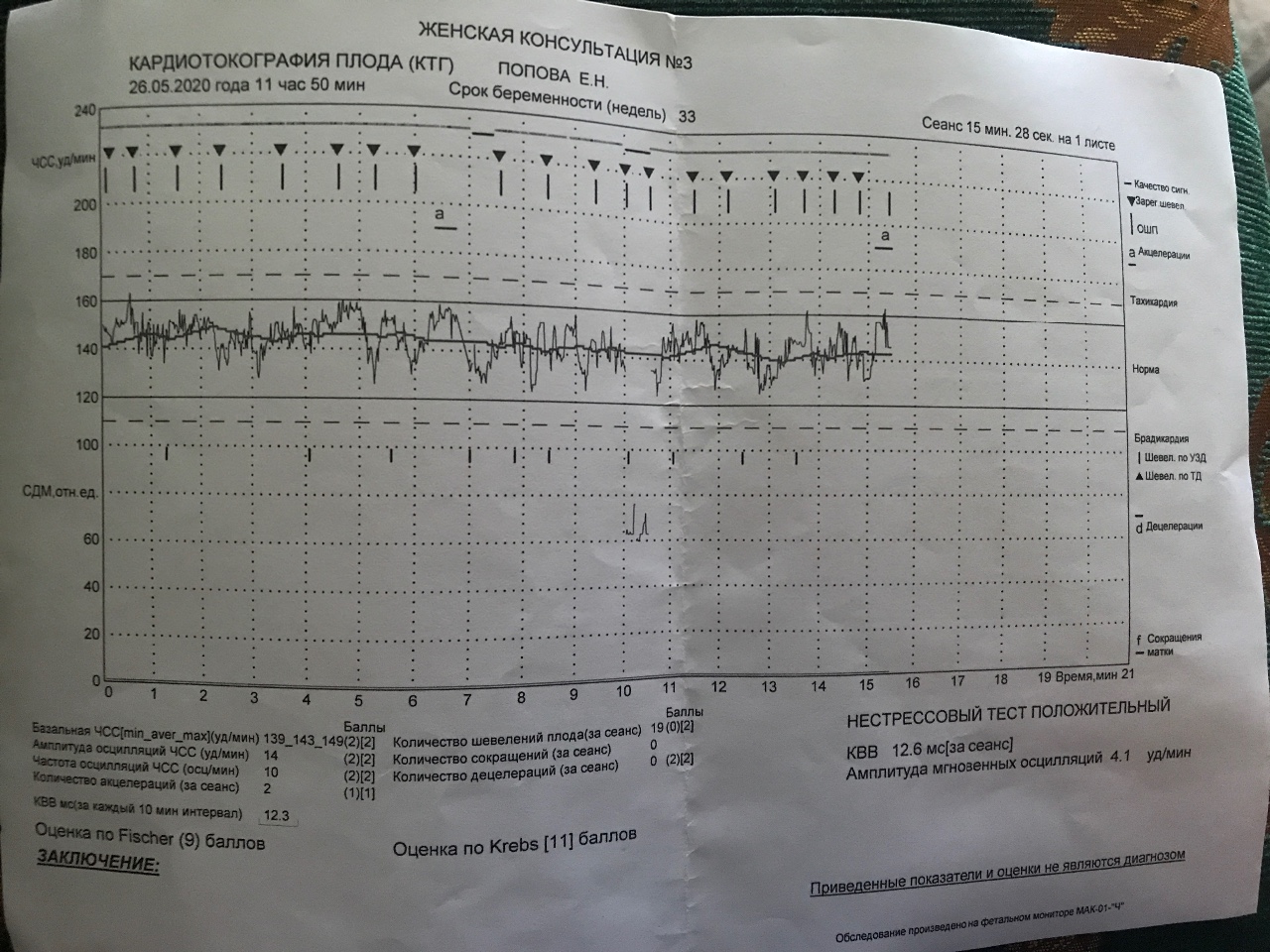
Самым важным вопросом этого метода остается расшифровка КТГ плода . Существует несколько шкал, самыми популярными считаются 10-бальная шкала Фишера и 12-бальная шкала Кребса. Обычно оценивают показатели по обеим шкалам и пишут две оценки в заключении. Стоит учитывать, что данные не должны расходиться по разным шкалам больше чем на три пункта.
Ниже остановимся подробнее на шкале Фишера. После проведения КТГ расшифровка результатов идет по следующим параметрам, которые по отдельности оцениваются от 0 до 2 баллов:
1. Базальный ритм (среднее между значениями сердцебиения плода, не меняющаяся на протяжении 10 минут и дольше):
- меньше 100 или больше 180 ударов в минуту – 0 баллов,
- от 100 до 119 ударов в минуту и от 161 до 180 ударов в минуту – 1 балл,
- от 120 до 160 ударов в минуту – 2 балла.
2. Вариабельность (амплитуда):
- меньше 3 ударов в минуту – 0 баллов,
- от 3 до 5 ударов в минуту – 1 балл,
- от 6 до 25 ударов в минуту – 2 балла.
3. Вариабельность (частота в 1 минуту):
- меньше 3 – 0 баллов,
- от 3 до 5 – 1 балл,
- больше 6 – 2 балла.
4. Акселерации (повышение частоты сердечных сокращений плода на 15-20 ударов в минуту относительно базовой частоты, возникающее в ответ на шевеление плода, прижатие пуповины, сокращение матки) за 30 минут:
- 0 (отсутствие) – 0 баллов,
- 1-4 (периодические) – 1 балл,
- от 5 и выше (спорадические) – 2 балла.
5. Децелерации (снижение частоты сердечных сокращений в ответ на шевеление или сокращение матки) за 30 минут:
- тяжелые атипичные децелерации – 0 баллов,
- легкие среднетяжелые децелерации – 1 балл,
- отсутствие или короткие неглубокие децелерации – 2 балла.
Чтобы избежать субъективности при расшифровке данных КТГ плода, в современном мире медицины стараются создавать приборы и компьютерные программы, которые максимально автоматизируют процесс расшифровки.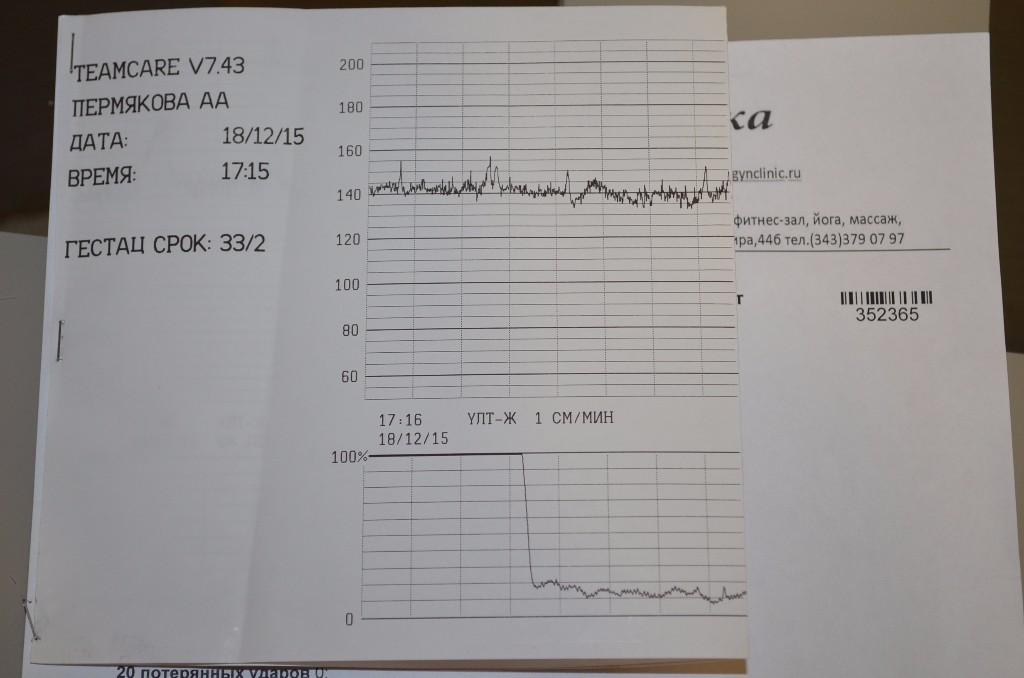
В норме КТГ при беременности по шкале Фишера составляет диапазон данных от 8 до 10 баллов. Результат в 6-7 баллов считается предпатологическим, и врачи, скорее всего, назначат повторное обследование. Если результаты КТГ меньше 6 баллов, это, скорее всего, означает внутриутробную гипоксию плода и требует немедленной госпитализации или срочного родоразрешения.
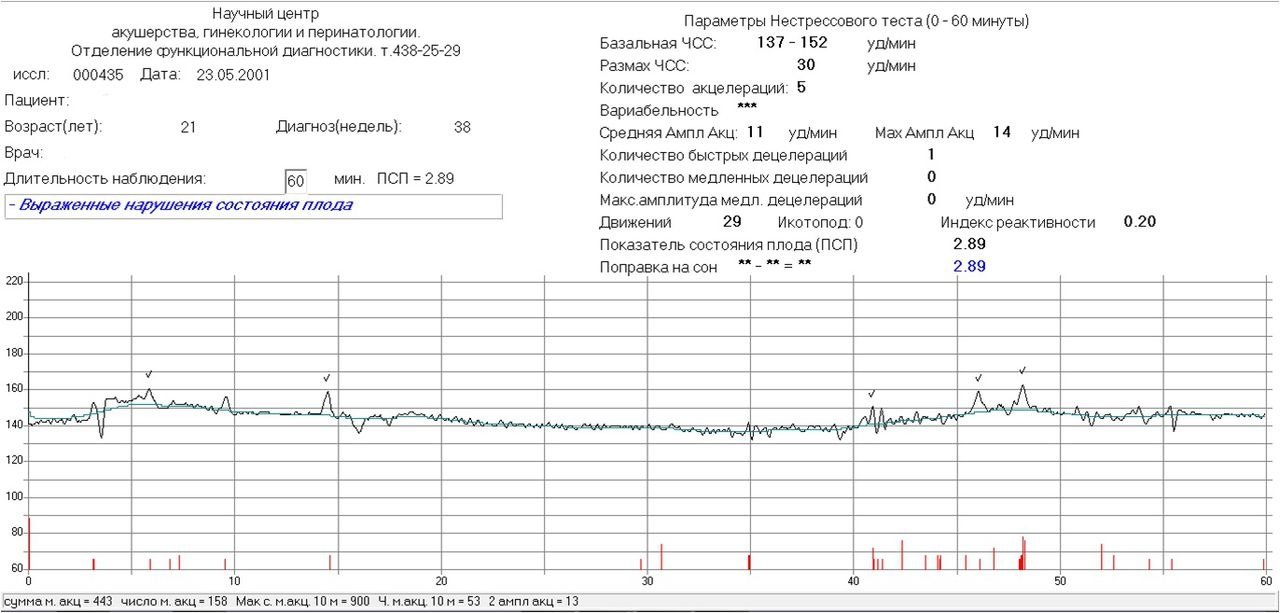
Показатель состояния плода (ПСП)
По результатам графика, полученного при КТГ, врачи находят значение ПСП (показателей состояния плода), которое при нормальном развитии составляет меньше 1. Если эти значения находятся в диапазоне от 1 до 2, это может свидетельствовать о начале появления нарушений у плода. Если значение ПСП больше 3, это говорит о критическом состоянии плода. Однако на основании только этих данных никаких решений не принимается, рассматривается вся история протекания беременности. Причинами отклонения показателей могут быть не только проблемы в развитии плода (сердечная недостаточность, анемия, гипоксия), но и некоторые состояния у будущей матери и ребенка, не связанные с нарушениями (повышенная температура у беременной, фаза сна у ребенка).
Стоит отметить, что метод КТГ является вспомогательным или дополнительным, поэтому на его результаты опираются только в совокупности с другими диагностическими данными. Главным образом, это влияет на небольшие отклонения от нормы, поэтому бить тревогу в случае диагностики результатов, отличных от нормы, перед разговором и обсуждением результатов с гинекологом не стоит.
4.2
Приблизительное время чтения: 8 минут
Наряду с многочисленными анализами и исследованиями женщинам назначается такая важная процедура, как кардиотокография. КТГ плода при беременности назначается всем женщинам без исключения для диагностики состояния малыша и определения степени его развития. Как правило, процедура назначается несколько раз за весь период и помогает наблюдать беременность, протекающую с осложнениями и патологиями.
Что такое КТГ
Абсолютно безопасный и безболезненный метод оценки общего состояния малыша способом оценки частоты сердечного ритма. Также помогает оценить состояние матки и степень ее тонуса.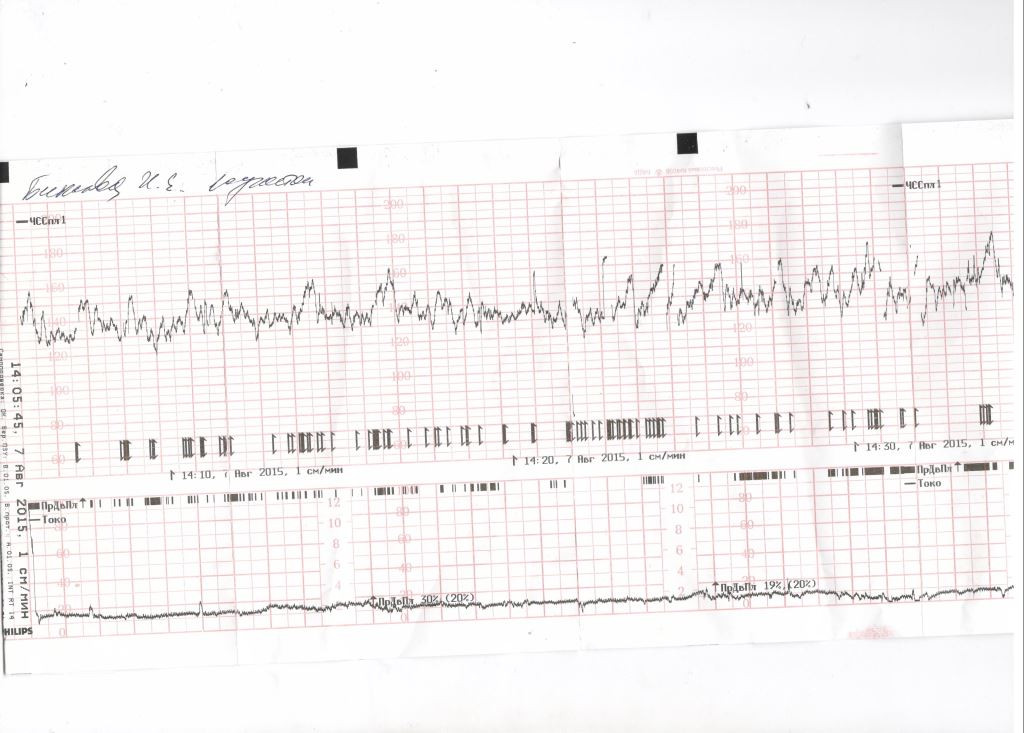 Полученные данные передаются на калиброванную ленту, где изображены графические сигналы. Процесс обследования проводится при помощи ультразвукового датчика, данные передаются на монитор компьютера, а затем оператором переносятся на ленту.
Полученные данные передаются на калиброванную ленту, где изображены графические сигналы. Процесс обследования проводится при помощи ультразвукового датчика, данные передаются на монитор компьютера, а затем оператором переносятся на ленту.
КГТ делится на два типа:
- Наружная процедура проводится при целостности плодного пузыря. При исследовании датчики крепятся к определенным местам живота, чтобы получить максимально глубокий и стабильный сигнал.
- Внутреннее исследование назначается при нарушении плодной оболочки, как правило, перед родами. Электрод помещается в полость матки, чтобы оценить внутриматочное давление, которое может помешать планомерному родоразрешению.
Данный метод исследования показывает высокую степень результативности, особенно при имеющихся патологиях. Современная интранатальная и антенатальная кардиотокография обязательно должна назначаться каждой будущей матери, чтобы иметь возможность контролировать состояние младенца, определять тактику ведения беременности и принимать решение о способе родоразрешения.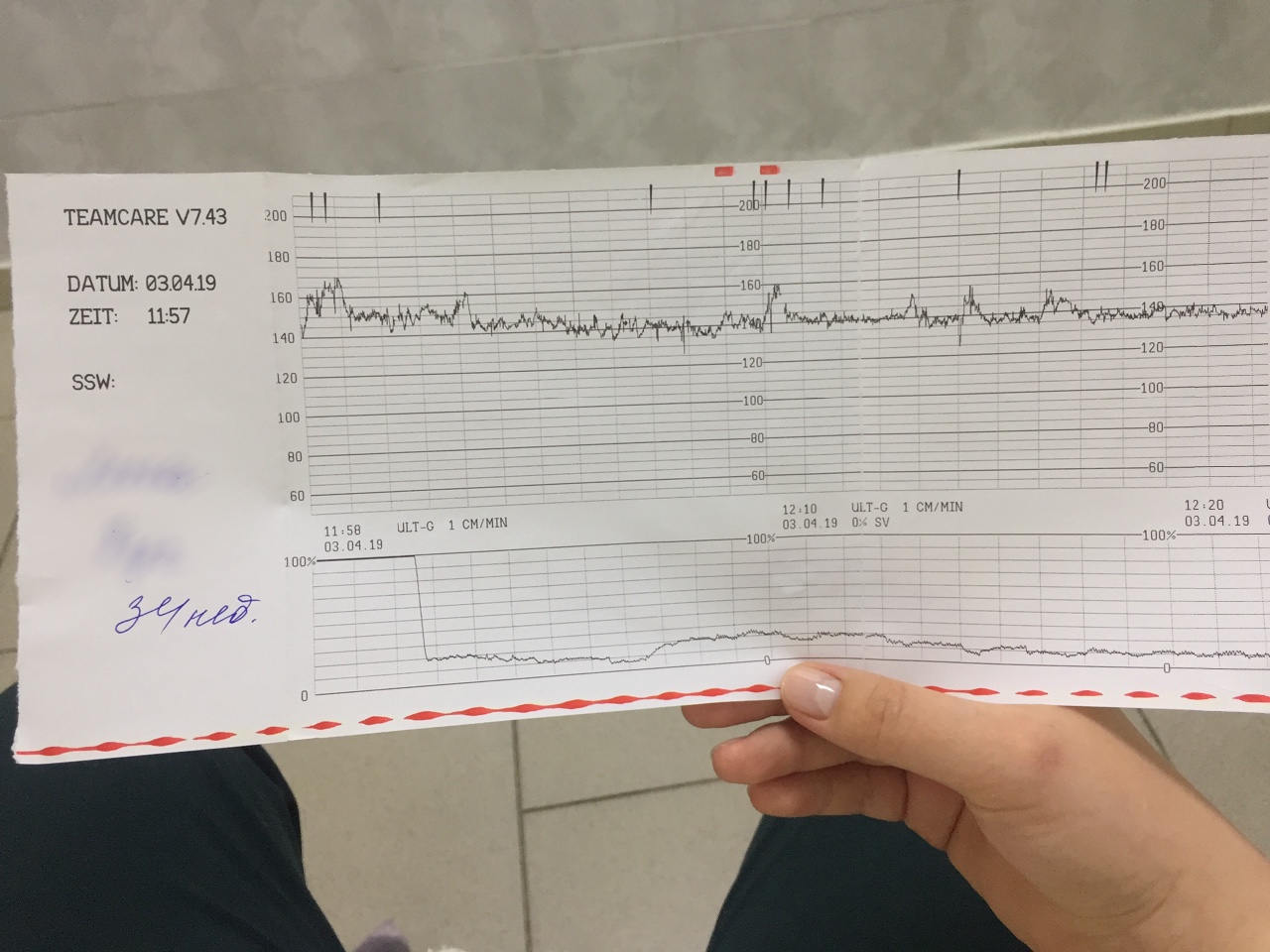
Плановый календарь КТГ
Многие будущие мамы интересуются, что представляет собой КТГ, на каком сроке делают , и каких результатов ожидать? Стоит отметить, что на плановое исследование женщины направляются с 28 по 32 неделю . Если наблюдаются отклонения от нормы по общим анализам, КТГ обследование может быть назначено гораздо раньше планируемого срока.
Если диагностика проводилась на ранних сроках, около 28 недель , тогда в обязательном порядке КТГ назначается на 33-34 неделе. При отсутствии патологических нарушений исследование проводится на сроке 30-32 недели, а затем перед запланированными родами. В зависимости от имеющихся осложнений, Минздрав рекомендует проводить исследование с 28 недели, по такому графику:
- Если беременность переношенная – каждые 5 дней после планируемого срока родов.
- Многоводная беременность – не реже 1 раза в 7 дней.
- При несовместимой группе крови – каждые 14 дней.
- Если были обнаружены пороки развития – еженедельно.
- Если малыш крупный или мама перенесла инфекционные болезни на первом триместре – каждую декаду месяца.
- Будущие мамочки старше 35 лет – каждую декаду.

Многие интересуются, как часто делают КТГ при беременности и почему каждый гинеколог решает самостоятельно, когда проводить исследование. Зачастую курирующий врач руководствуется своим опытом и знаниями, а также назначает исследование, исходя из полученных диагностических данных других обследований. Опытные специалисты отправляют на КТГ по такому принципу:
- Благоприятно протекающая беременность – КТГ после восьмого месяца можно делать каждые 14 дней.
- При отягощенном фоне – каждую неделю с седьмого месяца.
- При негативных результатах прошлого обследования проводится дополнительное КТГ и при неутешительных результатах беременная направляется в стационар.
Исключение составляют те беременные, которым не была назначена кардиотокография. В такую категорию вошли женщины с отличным состоянием здоровья и отсутствием каких-либо подозрений на патологии.
Показания к проведению кардиотокографии
Обследование при помощи ультразвука назначается таким категориям беременных:
Каждая женщина, подпадающая под любую категорию, имеет степень риска развития пороков плода, поэтому обязана пройти высокоточную диагностику, чтобы оценить степень развития малыша и исключить негативные последствия.
Техника проведения КТГ
Достаточно актуальный вопрос, который часто слышат гинекологи – это как делают КТГ беременным и нужно ли ожидать от процедуры неприятных ощущений? Нужно успокоить будущих мам, которым выдано направление на кардиотокографию: никаких неприятных ощущений или дискомфорта не будет, поэтому нужно максимально успокоиться, чтобы получить достоверные результаты.
Процесс проводится на кушетке, где беременная размещается так, как удобно именно ей. Далее по нижней и боковым частям живота крепятся специальные сверхчувствительные датчики, передающие полученную информацию на монитор специалиста. Для диагностики используется два типа датчиков:
Датчики для проведения КГТ для беременных
- Датчик давления (тензодатчик) — оценивает критерии матки, ее тонус и частоту сокращений.
- Ультразвуковой – улавливает частоту и ритм сердечной деятельности малыша.
Сама процедура длится около 60 минут, иногда меньше. Все зависит от того, насколько активен малыш в это время. Чтобы сократить время процедуры и подбодрить кроху к «активным действиям», можно съесть за час до процедуры шоколадку. Также нужно иметь с собой питьевую воду и небольшой перекус в виде яблока.
Подготовка к КТГ при беременности минимальная и не требует от женщины каких-либо ограничений в питании или двигательной активности. Единственное, что требуется от будущей мамы, – это минимум стресса и переживаний накануне исследования, так как можно получить погрешности результата. Женщине нужно успокоить себя и понять, что если до этого срока не было выявлено никаких патологических отклонений, то и беспокоится не о чем.
Результаты КТГ – расшифровка и нормы
Полученные графические результаты должен считывать только лечащий врач, который по своему опыту может видеть не только факты, но и замечать некоторые нестыковки показателей, полученные из-за слабой активности малыша. При разшифровке КГТ при беременности исследовании учитывают такие показатели:
При разшифровке КГТ при беременности исследовании учитывают такие показатели:
- Уменьшение и увеличение частоты сердечного ритма малыша в активную фазу и при сокращениях матки.
- Влияние сокращение матки на движение плода.
- Базальный и вариабельный ритм сердца ребенка.
Интерпретировать полученные данные не сложно, и медики применяют методику, где каждому показателю дается балльная оценка. К примеру, что показывает КТГ при беременности в цифрах:
- 2 – хорошие показатели;
- 1 – наблюдается начальная стадия патологии;
- 0 – явно выраженные признаки патологии.
Базальный ритм в норме составляет 110-170 ударов за минуту. Такие показатели фиксируются числом 2. Если есть легкие нарушения, тогда нормальные показатели повышаются или понижаются на 10 делений. При угрожающих показателях ритм будет составлять менее 100 ударов или более 180 за минуту.
Вариабельный ритм оценивается по частоте амплитуды и осцилляции.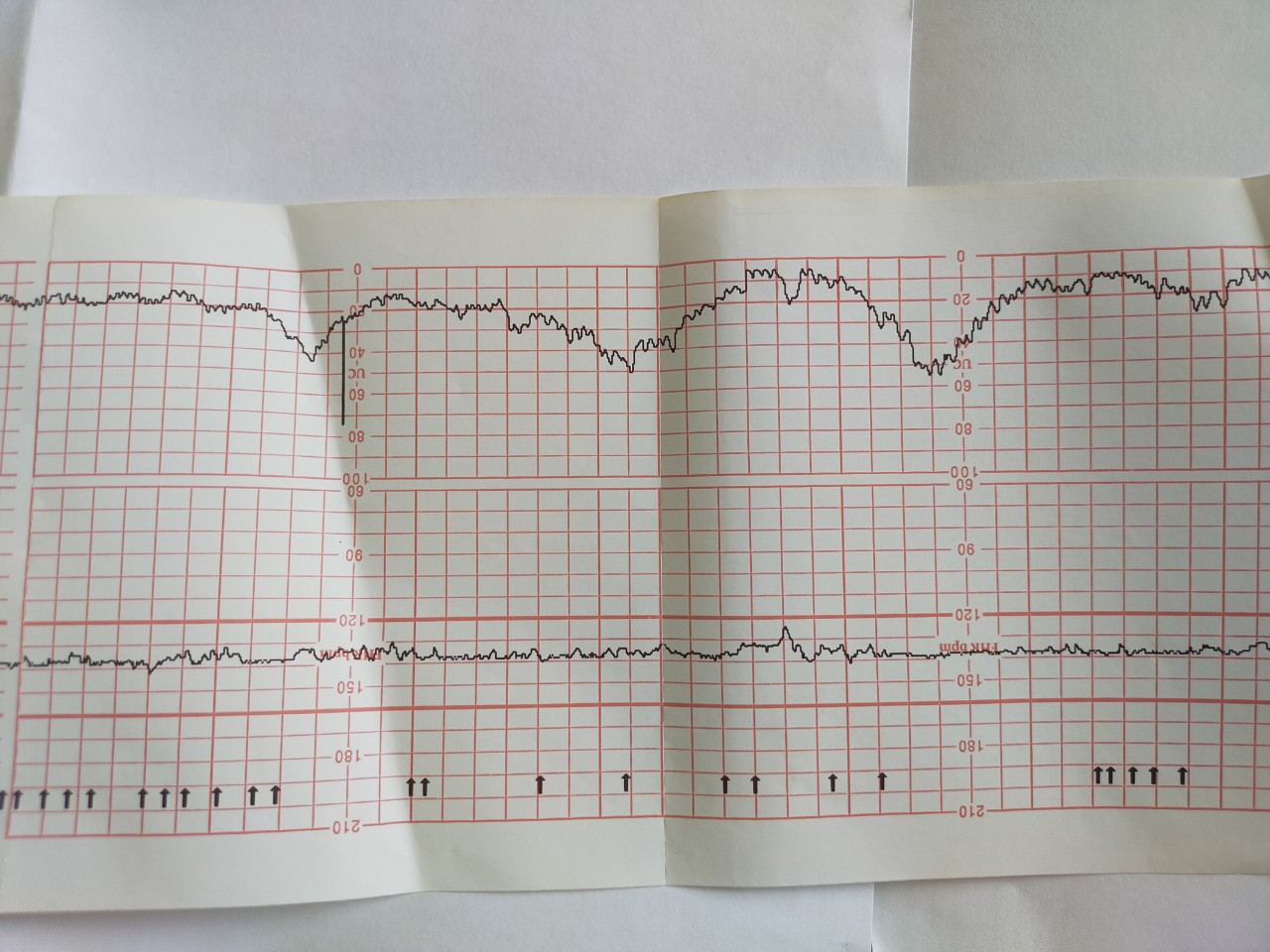 Данные исходят из активности плода и сокращения матки относительно базального ритма. Нормой при вариабельном ритме является 10-25 ударов за минуту. Насторожить должны результаты 5-9 или более 25 ударов за минуту. Если же показатели фиксируют менее пяти ударов в минуту или более 25 – необходима госпитализация.
Данные исходят из активности плода и сокращения матки относительно базального ритма. Нормой при вариабельном ритме является 10-25 ударов за минуту. Насторожить должны результаты 5-9 или более 25 ударов за минуту. Если же показатели фиксируют менее пяти ударов в минуту или более 25 – необходима госпитализация.
Акселерация – степень частоты биологического ритма организма за 30 минут. Показатель более 5 – норма, от 4 до 1 – допустимо, не просматривается ни одного ритма – нарушение развития.
Децелерация – снижение частоты ритма. Если за первые 10 минут была зарегистрирована такая частота, тогда ставится отметка норма – 2 балла. Ритм зафиксирован через 20 минут – 1 балл, при отсутствии – 0 баллов.
- 10-8 баллов – результаты КТГ плода норма, они указывают на здоровье малыша и благоприятный исход родов;
- 7-5 число указывает о легкой патологии, которую нужно наблюдать и корректировать;
- 4 и менее свидетельствуют о том, что необходимо срочно проводить кесарево сечение или стимулировать родовую деятельность.
Проведенное обследование на 32 или 35 неделе не имеет особых изменений, разве что в частоте базального ритма.
| Критерии | 0 баллов | 1 балл | 2 балла |
|---|---|---|---|
| Базальный ритм, уд/мин | меньше 100 или больше 180 | 100-120 или 160-180 | 121-159 |
| Вариабельность, амплитуда, уд/мин | меньше 3 | 3-5 | 6-25 |
| Вариабельность, число в минуту | меньше 3 | 3-6 | больше 6 |
| Число акцелераций за 30 минут | нет | 1-4 единичные или периодические | больше 5 единичных |
| Децелерации | поздние или вариабельные тяжелые, атипические | ранние (тяжелые) или вариабельные (легкие, умеренные) | отсутствуют или ранние (легкие, умеренные) |
| Шевеление плода | нет | 1-2 | больше 3 |
Наносит ли вред плоду КГТ и могут ли быть погрешности результата?
Ультразвуковые волны, которые используются при проведении процедуры, имеют самые низкие частоты, поэтому не оказывают никакой негативной реакции на малыша и его мать. За многолетнюю практику проведения КТГ не было зафиксировано ни одного случая, который бы указал на развитие патологии или отклонение развития. Поэтому беспокоиться о том, вредно ли КТГ для плода, не стоит, так как достоверных сведений на этот счет нет.
За многолетнюю практику проведения КТГ не было зафиксировано ни одного случая, который бы указал на развитие патологии или отклонение развития. Поэтому беспокоиться о том, вредно ли КТГ для плода, не стоит, так как достоверных сведений на этот счет нет.
Иногда женщины отмечают, что на момент исследования малыш начинает вести себя активно или наоборот затихает. Такая реакция связана со звуком, который подается малышу, так как датчики устанавливаются достаточно сильно.
Многих беременных женщин интересует вопрос, дает ли погрешность прослушивание и какие факторы могут повлиять на негативный результат. Безусловно, такой факт существует, и причиной тому такие моменты:
- Если беременная обильно поела перед процедурой.
- Запись была сделана в спокойную фазу, когда ребенок спал.
- Если женщина имеет любую стадию ожирения, можно ожидать погрешности в результатах по причине плохой передачи ритма через жировую клетчатку.
- Запись будет с погрешностями, если ребенок чрезмерно подвижен.
- При многоплодной беременности услышать все сердечки проблематично.

Какие патологии поможет выявить КТГ
Компьютерная томография плода максимально информативна, и дает возможность распознать тяжелые патологии и осложнения, которые влекут серьезные последствия для жизни ребенка. При использовании данного метода,в медицинской практике удалось снизить критические показатели внутриутробной смертности на 25-45% в зависимости от региона. Определяя наличие серьезных отклонений, гинекологам удается вовремя повлиять на ситуацию, и сохранить здоровье матери и ребенка.
Фетоплацентарная недостаточность – часто встречающаяся патология, которая в 50% несет гибель плода. Зачастую причиной становится поздний гестоз, бактериальные или вирусные инфекции матери. Чтобы понимать, зачем делают КТГ беременным, необходимо оценивать всю степень тяжести, которую могут принести такие аномалии, как ФПН.
Внутриутробные инфекции – это целая группа заболеваний, которые несут массу патологических изменений, врожденных аномалий, задержку умственного и физического развития, и гибели плода. При ранней диагностике удастся максимально снизить токсичность инфекций и устранить патологические явления.
При ранней диагностике удастся максимально снизить токсичность инфекций и устранить патологические явления.
Раннее старение плаценты грозит развитием гипоксии и голоданием плода , который недополучает питательные элементы и витамины. Если ребенок будет испытывать кислородное и питательное голодание, это отразиться на психоэмоциональной, сердечно-сосудистой и пищеварительной системах.
Гипоксия – внутриутробный синдром кислородного голодания, который может стать причиной отека мозга у младенца, привести судорогам, пневмонии, некрозу, асфиксии. Кислородная недостаточность требует стационарного лечения, чтобы нормализовать фетоплацентарное кровообращение.
Гестационный диабет беременных может стать причиной асфиксии плода, и привести к гипергликемии, когда организм малыша не способен самостоятельно вырабатывать инсулин. Стоит сказать, что данный недуг требует длительного и внимательного лечения даже после родов, так как в большинстве случаев, у женщин в дальнейшем вырабатывался сахарный диабет.
Кардиотокография при беременности (КТГ)
КТГ (кардиотокография) – это метод исследования сердцебиения плода и маточных сокращений у беременных, при котором все данные записи фиксируют на специальную ленту. Показатели ЧСС у ребенка будут зависеть от некоторых факторов, таких как: время суток, и от наличия факторов риска.
Как же расшифровываются конечные показатели КТГ?
Расшифровка конечных , проводится специалистом с учетом таких данных, как: вариабельность ЧСС плода, базальный ритм, акцелерация, децелерация и двигательная активность плода. Такие показатели, в конце обследования, отображаются на ленте, и имеют вид графиков, разных по форме. Итак, рассмотрим подробнее выше перечисленные показатели:
- Вариабельностью (или амплитудой) называют нарушения частоты и регулярности сократительных движений ритма и амплитуды сердца, которые основаны на полученных результатах базального ритма. Если не наблюдается никакой патологии развития плода, показатели ЧСС не должны быть однообразны, это хорошо видно с помощью визуализации постоянной сменой числовых показателей на мониторе, во время обследования КТГ. Изменения в пределах нормы могут колебаться в диапазоне 5-30 ударов в минуту.
- Базальным ритмом обозначается средние показатели ЧСС малыша. Показателями нормы является биение сердца от 110 и до 160 ударов в одну минуту во время спокойствия плода и женщины. Если ребенок активно шевелится, ЧСС будет держаться от 130 до 180 ударов в продолжении одной минуты. Показатели базального ритма в пределах нормы означают отсутствие гипоксического состояния плода. В случаях, если показатели ниже нормы или выше, считается, что присутствует гипоксическое состояние, которое отрицательно сказывается на нервной системе малыша, которая находится в недоразвитом состоянии.
- Под акцелерацией подразумевают учащенное количество сердцебиения, сравнительно уровня показателей базального ритма. Показатели акцелерации воспроизводятся на кардиотокограмме в виде зубчиков, норма составляет от двух-трех раз за 10-20 минут. Возможно не большое увеличение учащенности до четырех раз за 30-40 минут. Патологией считается, если акцелерация и вовсе отсутствует за период в 30-40 минут.
- Децелерация – это снижение частоты сердечных ударов сравнительно со степенью базальных показателей сердечного ритма. Показатели децелерации имеют форму провалов или по-другому отрицательных зубьев. В пределах нормального функционирования плода, данные показатели должны полностью отсутствовать или совсем незначительно проявляться по глубине и длительности, и очень редко встречаться. После 20-30 минут КТГ исследования при проявлении децелерации, возникают подозрения в ухудшении состояния будущего малыша. Большое беспокойство в развитии плода вызывают повторное и разнообразное проявление децелерации во время всего обследования. Это может стать сигналом о наличии декомпенсированного стресса у плода.
- Вариабельностью (или амплитудой) называют нарушения частоты и регулярности сократительных движений ритма и амплитуды сердца, которые основаны на полученных результатах базального ритма. Если не наблюдается никакой патологии развития плода, показатели ЧСС не должны быть однообразны, это хорошо видно с помощью визуализации постоянной сменой числовых показателей на мониторе, во время обследования КТГ.
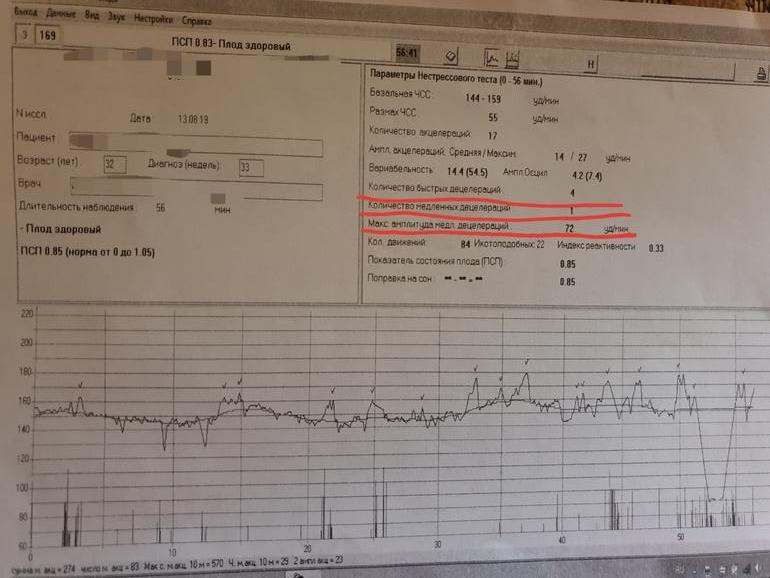 Изменения в пределах нормы могут колебаться в диапазоне 5-30 ударов в минуту.
Изменения в пределах нормы могут колебаться в диапазоне 5-30 ударов в минуту.
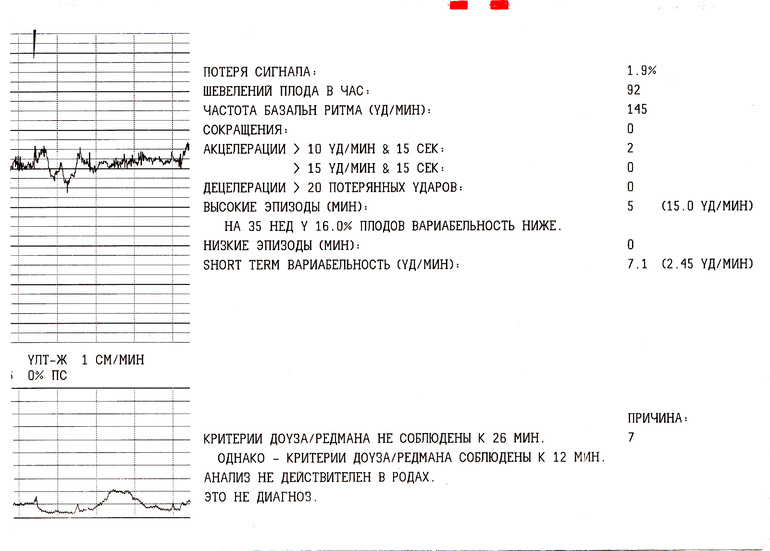
Важность показателей состояния плода (ПСП)
После того, как готовы графические результаты КТГ исследования, специалист определяет значение показателей состояния плода. Для нормального развития ребенка эти значения будут меньше 1. Когда показатели ПСП составляют от одного до двух, это говорит о том, что состояние плода начинает ухудшаться и появляются некоторые не благоприятные изменения.
Когда показатели ПСП свыше трех, это значит, что плод находится в критическом состоянии. Но при наличие только таких данных специалист не может принимать никаких решений, сначала будет рассматриваться полностью анамнез течения беременности.
Нужно понимать, что не только патологические процессы в развитии малыша могут стать причиной отклонения показателей от нормы, это могут быть также некоторые состояния беременной и малыша, которые не зависят от нарушений (например, повышенные температурные показатели у беременной женщины или, если малыш находится в состоянии сна).
Какие баллы КТГ считаются нормой при проведении КТГ, считается патологией?
Результаты кардиотокографии оцениваются по специальной бальной шкале Фишера — присвоение 0-2 баллов к каждому из вышеперечисленных показателей. Затем баллы суммируются и делается общий вывод о наличии или отсутствии патологических изменений . Результат заявляет о неблагоприятном прогнозе – развитие гипоксии у плода, 6 бальное значение может говорить о начинающейся кислородной недостаточности.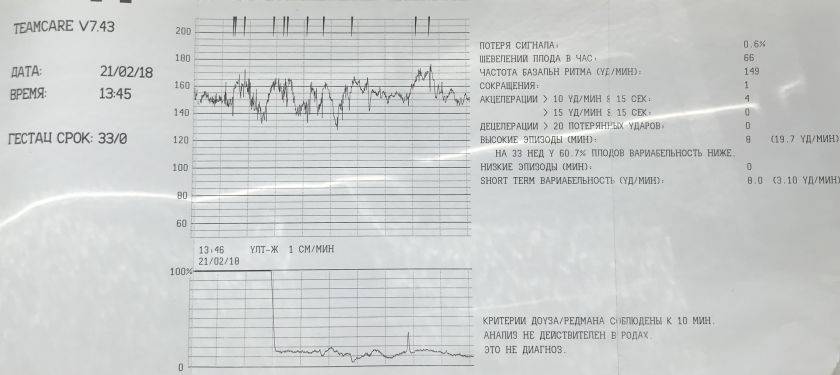
Что значит оценка КТГ 7 баллов в заключении?
КТГ 7 баллов – такая оценка считается показателем начала кислородной недостаточности плода. В данном состоянии специалист назначает соответственное лечение, чтобы избежать возникновение гипоксии, а также для улучшения состояния малыша при ее наличии. При оценке в 7 баллов на 32 неделе лечебные мероприятия начинают проводить без замедления. Врач, который наблюдает за течением беременности, может в срочном порядке, отправить женщину на стационарное лечение или ограничится капельницами на дневном стационаре.
Во время облегченной стадии голодания кислородом, обходятся более частым и длительным пребыванием на свежем воздухе, если позволяет погода. Или приемом медикаментозных средств для профилактики данного состояния.
Даже если после расшифровки КТГ обследования специалист определяет результат в 7 баллов, которые являются тревожным знаком, не стоит паниковать, ведь современная медицина сможет помочь будущему малышу избавиться от данного состояния.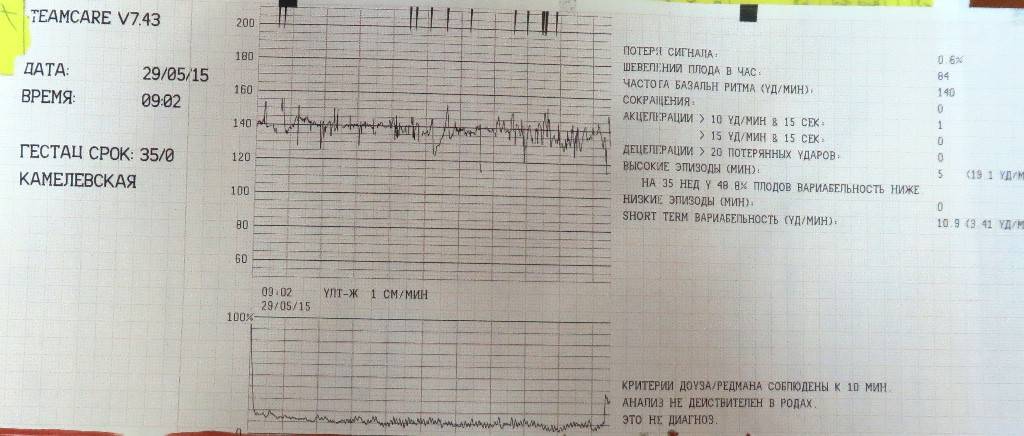
При выявленных патологических процессах в малыша, которые являются реакцией на сокращения матки, необходимо срочно с результатами исследования, обратиться к врачу-гинекологу. После оценки результатов специалист сможет назначить грамотное лечение, а также отправить на повторное прохождение КТГ.
Значение оценки КТГ 8 баллов
Очень многих будущих мамочек интересует вопрос 8 бального значения КТГ, являются ли данные показатели причиной для беспокойства? КТГ 8 баллов показывает нижнюю границу нормы, и такое состояние плода обычно не требует ни проведения лечебных мероприятий, ни госпитализации.
Какое значение имеют оценки в 9 и 10 баллов?
Нормальными значениями считается оценка 9 и 10 баллов. Данные показатели могут означать одно, что развитие плода проходит хорошо, без развития патологий. Оценка в 10 баллов говорит о том, что состояние будущего малыша в пределах нормы.
Какие патологические процессы можно выявить при исследовании КТГ?
Как же воспринимать результаты КТГ? Полагаясь только на полученные данные КТГ, невозможно окончательно определиться с диагнозом, так как патологические отклонения от 10-ти бальной нормы могут быть временным состоянием в ответ на какой-либо внешний раздражитель.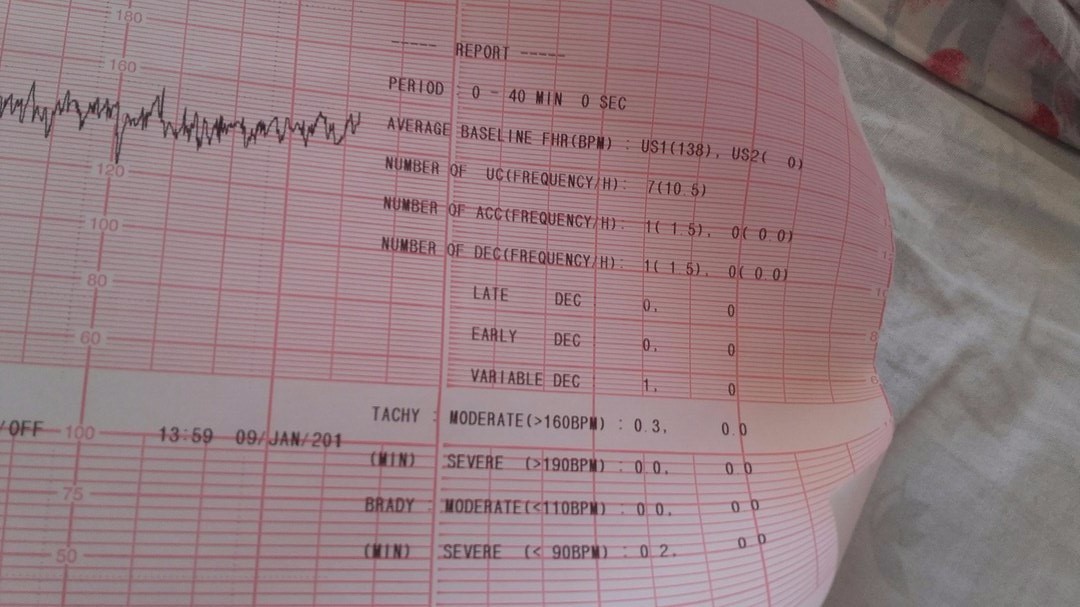 Данная методика проста в исполнении и поможет без особых затрат выявить отклонения от нормы в развитии плода.
Данная методика проста в исполнении и поможет без особых затрат выявить отклонения от нормы в развитии плода.
Метод КТГ поможет выявить следующие патологии:
Когда при расшифровке КТГ были выявлены отклонения от нормы, врач назначает УЗИ, а также . Если требуется, беременной расписывают лечение и повторное КТГ.
КТГ или кардиотокография – это безопасный метод функционального обследования во время беременности, помогающий врачам оценить, как плод ребенка усваивает кислород и справляется с физическими нагрузками, а именно, какая частота сердечных сокращений и их изменений в зависимости от движений плода, матери, а также сокращения самой матки.
Кардиотокография применяется не только во время беременности, но и во время родов, так как помогает врачам оценить состояние малыша во время прохождения по родовым путям. Чтобы лучше понять, что такое КТГ плода, зачем нужно это обследование, а также как расшифровать результаты, изучите данную статью.
Как делают КТГ плода?
Кардиотокография плода такая же важная как и УЗИ, и допплерометрия. Они не взаимозаменяемы, так как выполняют различного рода исследования.
Они не взаимозаменяемы, так как выполняют различного рода исследования.
Во время обследования методом КТГ сердечная деятельность плода фиксируется с помощью специальных ультразвуковых датчиков (1,5-2,0 Мгц), которые размешаются на животе беременной женщины в тех местах, где лучше всего прослушивается сердцебиение, поэтому перед этой процедурой проверяют сердечные тона с помощью стетоскопа. При этом беременная должна лежать либо на спине, либо на левом боку, возможно и положение сидя, при условии, что беременной так удобно.
Ультразвуковой датчик генерирует сигнал, который направляется к сердце плода ребенка, возвращается обратно и отображается на кардиомониторе в виде мгновенной частоты сердечных сокращений. Расчет ведет из количества ударов в 1 минуту. Все получаемые изменения фиксируются в системе в виде цифрового, светового и графического изображения.
Есть еще один датчик крепится в области дна матки для измерения ее сокращения и наблюдения за реакцией плода на эти сокращения. Исследование длится около 40-60 минут.
Исследование длится около 40-60 минут.
Очень важно, чтобы во время проведения обследования беременная чувствовала себя комфортно – удобно лежала, не нервничала. При этом нельзя, чтобы обследование проводилось натощак или сразу после еды. Дело в том, что любые изменения в уровне сахара в крови могут сказаться на результате обследования.
Зачем нужно делать КТГ плода?
Кроме того, что кардиотокография демонстрирует состояние сердца плода, являясь своеобразной расширенной кардиограммой еще не родившегося малыша, это обследование может выявить и много других отклонений в развитии. В первую очередь сюда необходимо отнести следующие патологии:
- Внутриутробная инфекция;
- Много- и ;
- Преждевременное созревание плаценты;
- Угроза преждевременных родов;
- Аномалии развития сердечно-сосудистой системы.
Но важно понимать, что КТГ плода не всегда дает 100% результат, так как бывают случаи, когда полученные данные недостоверны в силу некоторых причин. Например, бывает так, что ткани организма ребенка приспосабливаются к гипоксии, и аппарат не зафиксирует нехватку кислорода. Или наоборот, малыш может придавить головкой пуповину, мама понервничает перед обследованием или съест что-то не то, а аппарат выдаст «плохие» результаты. Поэтому, во-первых, очень важно выполнять процедуру обследования правильно, во-вторых, необходимо делать также дополнительные исследования, в том числе УЗИ, допплерометрию и т.д.
Например, бывает так, что ткани организма ребенка приспосабливаются к гипоксии, и аппарат не зафиксирует нехватку кислорода. Или наоборот, малыш может придавить головкой пуповину, мама понервничает перед обследованием или съест что-то не то, а аппарат выдаст «плохие» результаты. Поэтому, во-первых, очень важно выполнять процедуру обследования правильно, во-вторых, необходимо делать также дополнительные исследования, в том числе УЗИ, допплерометрию и т.д.
На каком сроке и как часто делают КТГ?
КТГ плода назначается не раньше 32 недели, потому что только к этому времени формируется взаимосвязь между двигательной активностью малыша и его сердечно-сосудистой системой, а также к этому времени уже работает установленный цикл сна и бодрствования ребенка. Период активности составляет приблизительно 50-60 минут, а покоя – 20-30. Во время обследования очень важно зафиксировать именно активную часть, когда малыш двигается. Если сделать КТГ раньше 32 недели, данные будут просто недостоверными.
При нормальном течении беременности КТГ плода делают максимум 1 раз в 10 дней. В случае выявления какой-то патологии, обследование проводится регулярно до тех пор, пока состояние не улучшиться.
Также КТГ делают и после отхождения околоплодных вод, при стимуляции родов и каждые 3 часа в первом периоде родов. Но как часто делать КТГ во время родов врач решает индивидуально, в зависимости от осложнений.
Расшифровка КТГ плода
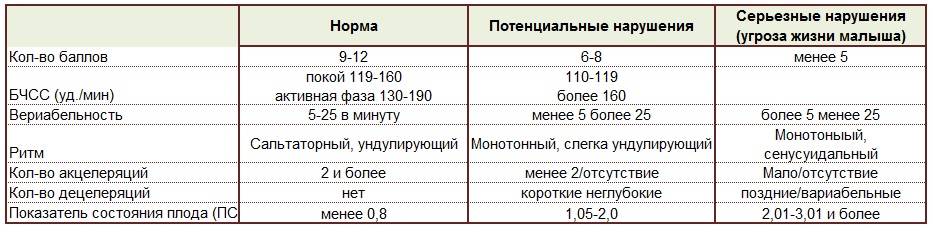
Результаты, полученные после обследования на кардиотокографе, расшифровывать должен врач. Полученные данные также как и в любом анализе или обследовании сравниваются с нормами. Но здесь не ставятся никаких диагнозов – так как обследование дает лишь дополнительные данные наряду с другими методами исследования. Кроме того, к получаемым данным нужно относиться осторожно, разумно сопоставляя с данными других обследований, а выполнить это профессионально может только врач. Но, если вы считаете, что ваш врач недостаточно доходчиво расшифровал полученные данные, воспользуйтесь таблицей.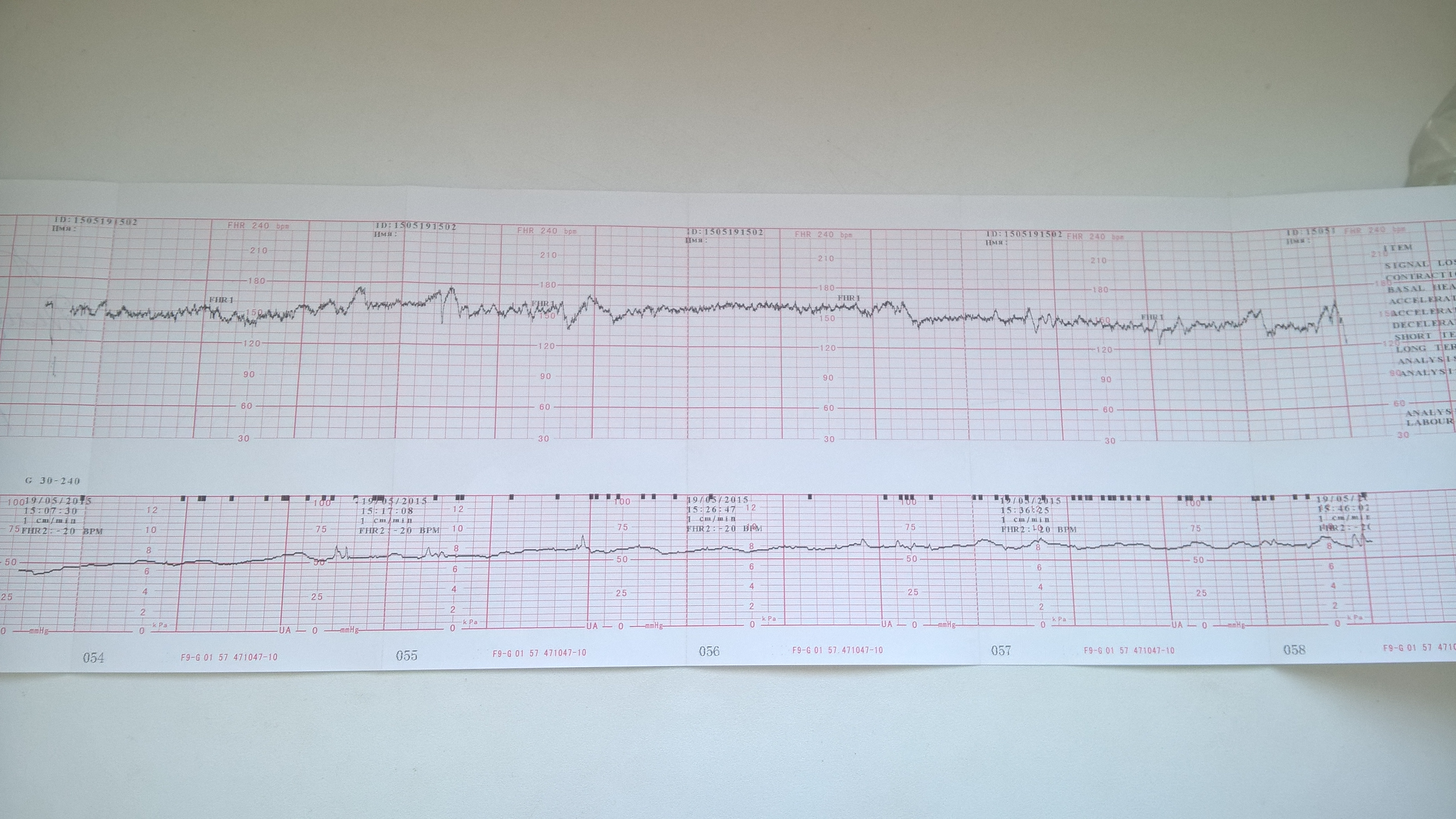
| Норма | Возможные нарушения | Серьезные нарушения (угроза для жизни ребенка) | |
| Количество баллов | 9-12 | 6-8 | 5 и менее |
| БЧСС (уд/мин) | 119-160 в покое, 130-190 — в активной фазе | 100-119 или больше 160 | 100 и меньше или больше 180 |
| Вериабельность | в пределах 5-25 в минуту | меньше 5 или больше 25 | меньше 5 или больше 25 |
| Ритм | ундулирующий или сальтаторный | монотонный или слегка ундулирующий | монотонный или синусоидальный |
| Количество акцелеряций | 2 и больше | меньше 2 или отсутствуют | мало или отсутствуют |
| Количество децелераций | нет | неглубокие и короткие | поздние и вериабельные |
| Показатель состояния плода (ПСП) | меньше 0,8 | 1,05-2,0 | 2,01-3,0 и выше |
Кардиотокография (КТГ) – это метод исследования, заключающийся в непрерывной регистрации частоты сердечных сокращений плода и сократительной активности беременной матки.
КТГ является информативным и безопасным методом исследования, помогая объективно оценивать состояние плода и в зависимости от этого выбирать тактику ведения беременной или роженицы. Кардиотокография быстро заняла лидирующее положение среди методов исследования состояния плода и вытеснила электро- и фонокардиографию.
Запись КТГ традиционно проводится в течение 40 минут, однако информативный результат получают, уже начиная с 15-20 минут записи. За это время можно оценить состояние плода, выявить признаки гипоксии и другие варианты неблагополучия.
Виды КТГ:
1) Непрямая или наружная КТГ. Непрямая КТГ широко используется во время беременности и I и II периодов родов.
2) Прямая или внутренняя КТГ.
Прямая КТГ производится во время родов при вскрытом плодном пузыре. Сердцебиение плода измеряется с помощью игольчатого электрода, вводимого в предлежащую часть плода. Запись маточных сокращений осуществляется с помощью катетера, вводимого в полость матки.
Прямая КТГ на сегодняшний момент практически не используется, так как является инвазивным методом исследования (то есть нарушающим целостность тканей). Это достаточно травматичный, трудоемкий и затратный метод исследования (сложность стерилизации тонкой техники). Поэтому все, о чем будет говориться далее, относится к непрямой КТГ.
Как выглядит КТГ?
Аппарат для КТГ выглядит как записывающее устройство с двумя датчиками. Крепление датчиков к животу производится эластичными ремнями.
Частота сердечных сокращений плода (сердцебиение) регистрируется ультразвуковым датчиком, устанавливаемым в месте наиболее четкого выслушивания сердечных тонов. Место выслушивания зависит от срока, положения (продольное, косое, поперечное) и предлежания (головное, тазовое). График этот называется тахограммой . По оси абсцисс отражается время в секундах. По оси ординат частота сердечных сокращений. На примере ниже это первый, самый верхний график.
Сократительная (мышечная) активность матки регистрируется тензометрическим датчиком (датчик, реагирующий на изменение давления, тонуса).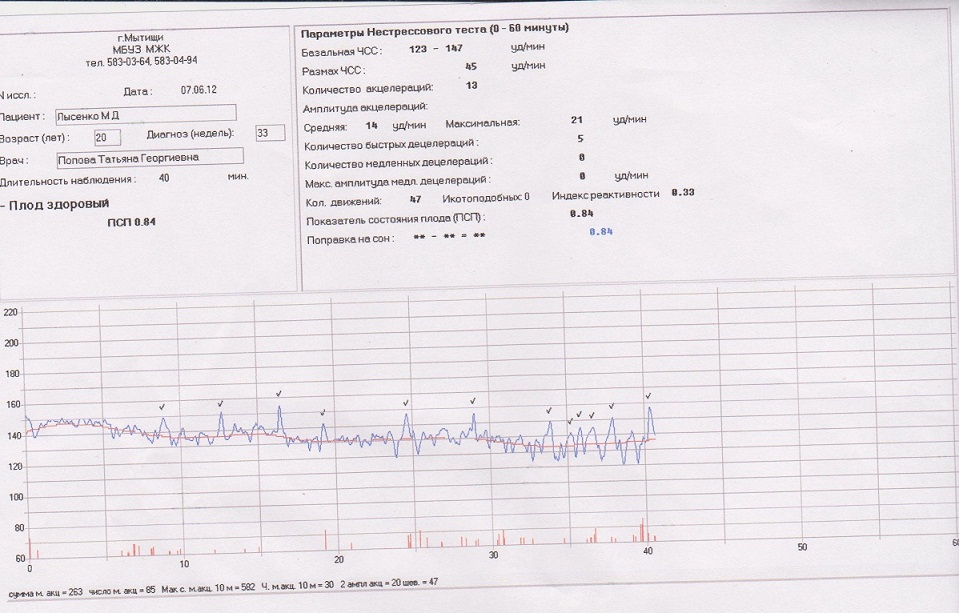 Датчик накладывается на правый угол матки, так как именно с этой области «запускается» волна сокращения матки. Этот график называется гистерограммой. На этой линии можно увидеть, что сокращений нет, либо есть периодический тонус, либо регистрируются схватки разной силы и регулярности. На примере ниже это третий график, самый нижний.
Датчик накладывается на правый угол матки, так как именно с этой области «запускается» волна сокращения матки. Этот график называется гистерограммой. На этой линии можно увидеть, что сокращений нет, либо есть периодический тонус, либо регистрируются схватки разной силы и регулярности. На примере ниже это третий график, самый нижний.
Также сейчас многие аппараты КТГ позволяют фиксировать шевеления плода. Это очень удобно и информативно, так как видно, как ребенок реагирует на собственные шевеления. На примере ниже это второй график, выделенный красным цветом.
Противопоказания КТГ
Противопоказаний к КТГ нет, процедура хорошо переносится пациентками и не влияет на ребенка, повторять КТГ можно столько раз. Сколько требуется для уточнения клинической ситуации.
Показания:
КТГ можно проводить примерно с 28 недель по особым показаниям, однако в сроке до 32 недель ребенок еще недостаточно зрелый и результат не будет максимально информативным.
Срок 28 недель не случаен, именно с этого срока формируется миокардиальный рефлекс, то есть реакция сердцебиения малыша в ответ на собственные движения. К 32 неделе этот рефлекс полностью сформирован и запись КТГ наиболее точно отражает его состояние.
К 32 неделе этот рефлекс полностью сформирован и запись КТГ наиболее точно отражает его состояние.
Плановое проведение КТГ
— беременность в сроке 32 недели и более
В III триместре выполняется 2-3 исследования КТГ. Если беременность протекает благополучно, то этого достаточно.
КТГ в родах
Запись КТГ проводится при поступлении в родзал всем женщинам, далее частота контроля зависит от клинической ситуации и определяется врачом.
Во II периоде родов ЧСС плода измеряется после каждой потуги, раньше это делали с помощью классического акушерского стетоскопа, сейчас используется датчик КТГ. Это информативно и удобно для врача и роженицы.
КТГ-контроль
Показаниями для более частого проведения исследования являются следующие ситуации:
— многоплодная беременность
Особенно трепетно контролируются монохориальные двойни, так как у малышей, имеющих общий «дом», то есть плодный пузырь, общее же питание и существует риск неравномерного распределения кислорода и питательных веществ между плодами.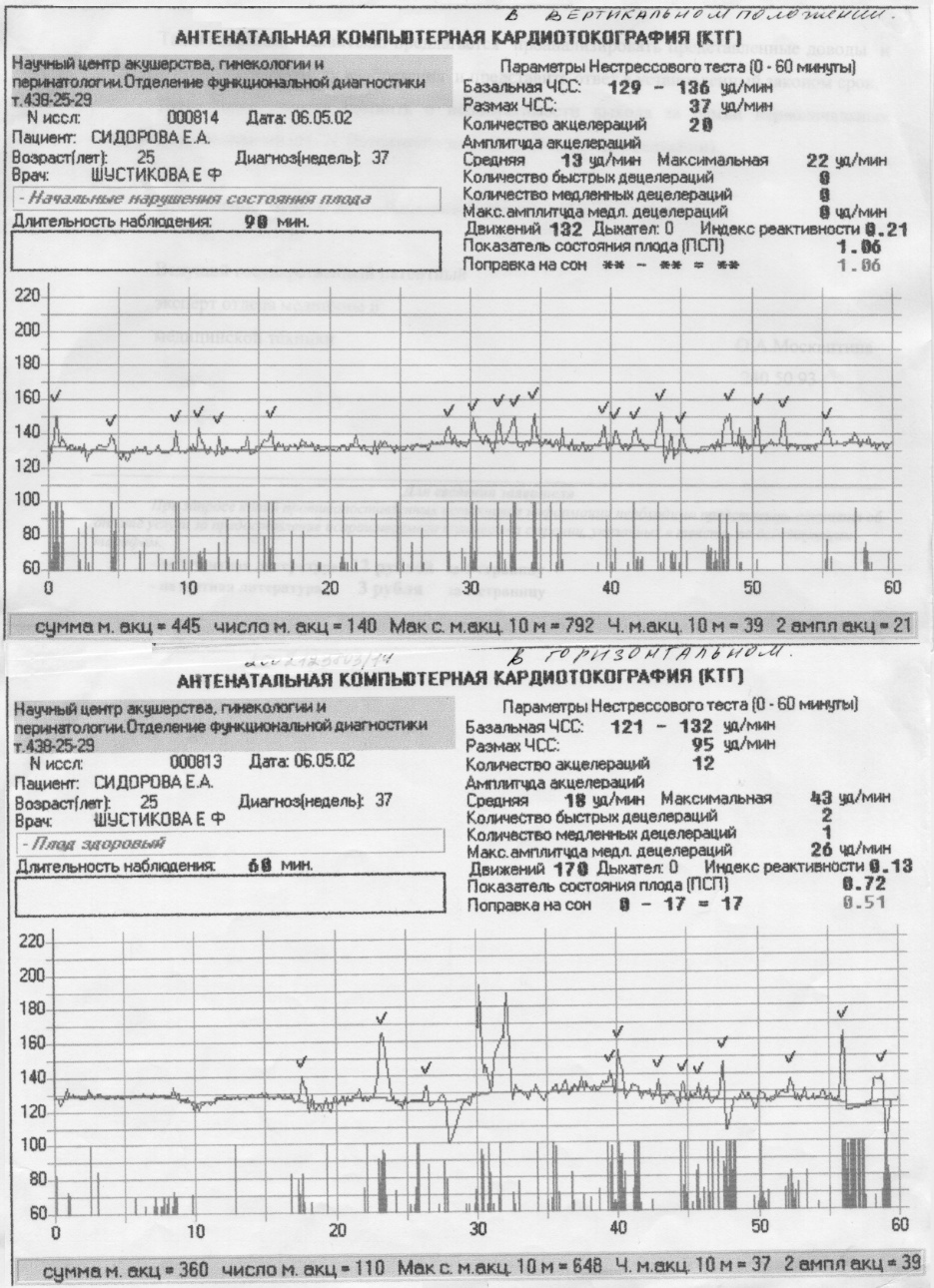 В этом случае один из малышей начинает испытывать кислородное голодание, как на приведенном ниже примере КТГ двойни.
В этом случае один из малышей начинает испытывать кислородное голодание, как на приведенном ниже примере КТГ двойни.
При маловодии у ребенка тоже выше риск гипоксии.
Многоводие может быть вызвано обменными нарушениями (самое главное из которых сахарный диабет), различными инфекциями и другими факторами, которые неблагоприятно сказываются на состоянии плода.
Переношенной считается беременность в сроке от 41 недели и 3х дней.
— снижение активности плода
При каждом посещении врач спрашивает, как вы ощущаете шевеления плода, активные или обычные, бурные или не ощущаете вообще. При наличии сомнений всегда спрашивайте своего доктора, нормальна ли та или иная ситуация.
— подозрение на патологию развития плода
КТГ здесь всегда сопутствует УЗИ и допплерометрии
— плацентарные нарушения
К этой группе патологий относятся: преждевременное старение плаценты, плацентарная недостаточность с гемодинамическими нарушениями (нарушение кровотока в маточных и пуповинной артериях) и без них. Такие нарушения могут возникать и на фоне полного здоровья женщины, однако чаще все же у курильщиц, женщин с сердечно-легочными заболеваниями и беременных в возрасте старше 35 лет.
Такие нарушения могут возникать и на фоне полного здоровья женщины, однако чаще все же у курильщиц, женщин с сердечно-легочными заболеваниями и беременных в возрасте старше 35 лет.
— отягощенный акушерский анамнез
Неблагополучное течение предыдущих беременностей и родов является фактором риска для данной беременности и поэтому требует повышенного внимания.
В родах КТГ более часто проводят женщинам, которым производится родовозбуждение или родоусиление окситоцином. Иногда датчик КТГ стоит весь период введения препарата и производится непрерывный мониторинг сердцебиения плода.
— ранее регистрировавшийся сомнительный тип КТГ
Сомнительный тип КТГ может быть обусловлен биоритмом сна, то есть в момент исследования ребенок спал, мало шевелился, и сердцебиение его мало менялось, график получился монотонным. Тогда следует провести контроль КТГ и должным образом подготовиться.
Как готовиться к КТГ?
Следует приходить не натощак, если в предыдущий раз на КТГ малыш спал, то сегодня съешьте что-нибудь сладкое, шевеления будут активнее;
Сходите в туалет, так как около 40 минут придется сидеть на одном местея;
Отключите телефон и прочие устройства, любые посторонние приборы могут вызвать помехи;
Перед процедурой немного подвигайтесь, пройдитесь, чтобы малыш не спал во время записи.
Как проводится КТГ?
Итак, вы пришли в кабинет КТГ…
Запись обычно проводится на кроватях или кушетках, следует занять положение на боку (обычно на левом) или полусидя, чтобы не было временного сдавления нижней полой вены беременной маткой.
Далее при помощи ремней вам прикрепят датчики, и вы будете находиться в этом положении от 15 до 40 минут, а возможно и дольше, если будет необходимость. Поэтому положение ваше должно быть еще и удобным для вас. Пить во время процедуры можно, поэтому можете поставить возле кровати бутылку с водой.
Как уже было сказано, сейчас большинство аппаратов автоматически регистрируют шевеления плода, но иногда этой функции нет, и тогда вам следует отмечать на выползающей из аппарата ленте периоды шевеления плода. Поинтересуйтесь у акушерки, которая ставит датчики, записываются ли здесь шевеления ребенка.
Расшифровка результатов КТГ
Анализ КТГ состоит из расшифровки ряда показателей:
1. Средняя частота сердечный сокращений
В норме ЧСС 120-160 ударов в минуту.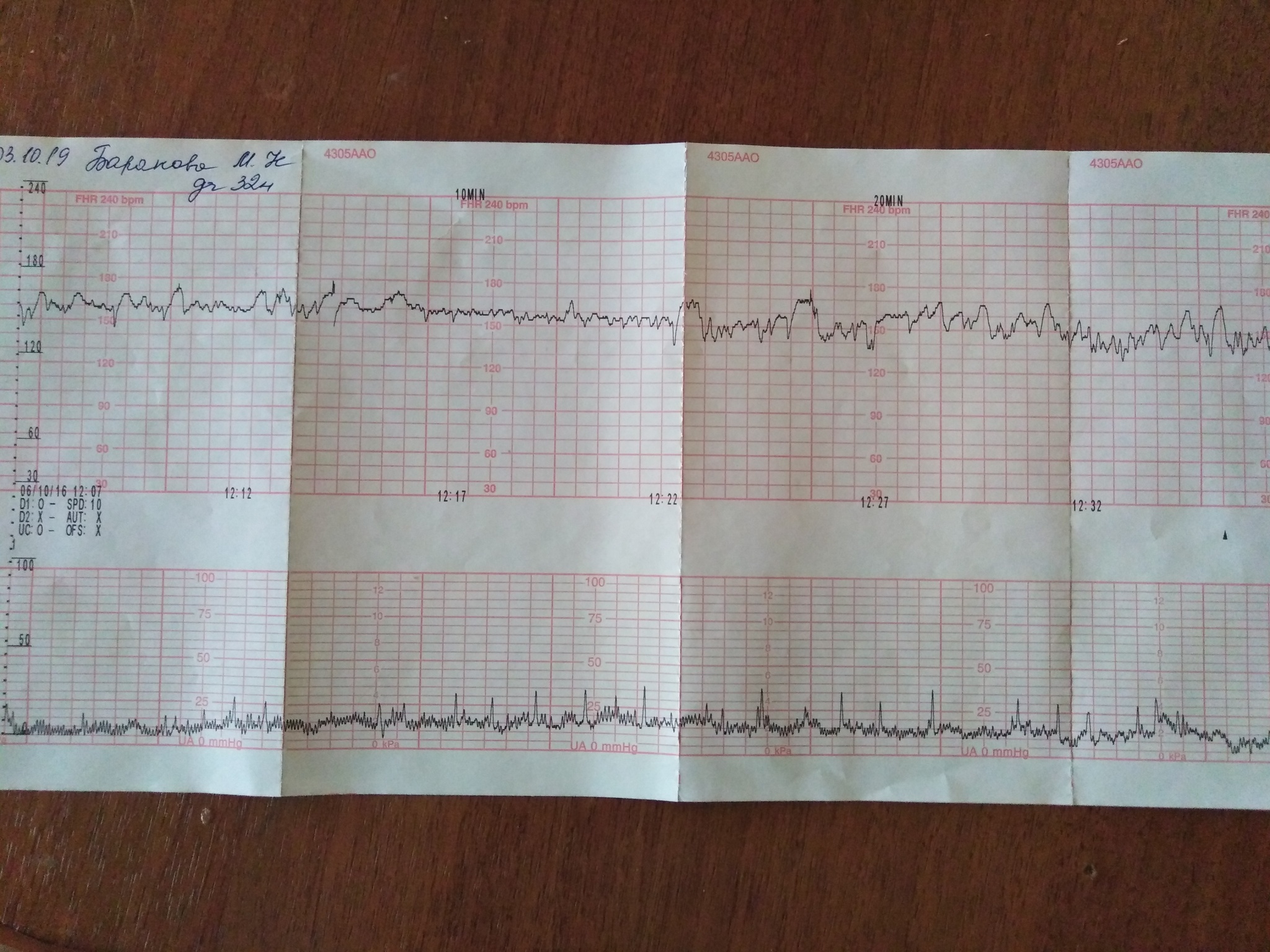 Брадикардия менее 100 и тахикардия более 180 в минуту – это серьезные признаки неблагополучия.
Брадикардия менее 100 и тахикардия более 180 в минуту – это серьезные признаки неблагополучия.
2. Вариабельность сердечного ритма
В норме амплитуда колебаний (отклонение от основной линии базальной частоты) 10-25 ударов в минуту и частотой более 6. То есть ритм «скачет», отклоняясь от базальной частоты и «скачкИ» (осцилляции) происходят около 6 раз в минуту.
Монотонный или немой ритм имеет амплитуду 0-5 в минуту
Слегка ундулирующий – 5-10 в минуту
Ундулирующий – 10-15 в минуту
Сальтаторный – 16-25 в минуту
В норме ритм ундулирующий или сальтаторный, либо указывается вариабельность 9-25 в минуту.
Характеристики «монотонный», «слегка ундулирующий» или «вариабельность ритма менее 9/ более 25 ударов в минуту» свидетельствуют о гипоксии плода.
3. Миокардиальный рефлекс
В ответ на собственные шевеления ЧСС ребенка увеличивается, и регистрируются акцелерации. Это называется нестрессовым тестом.
Стрессовый тест – это учащение ЧСС плода в ответ на раздражение сосков самой беременной, это связано с выбросом в кровь гормонов, повышающих тонус матки. Прочие стрессовые тесты, связанные с введением лекарственных препаратов, еще упоминаются в литературе, но сейчас не используются.
На примере ниже красным цветом отмечены шевеления плода и видно, что в ответ на активные шевеления появляются пики акцелераций на верхнем графике (тахограмме). Матка здесь находится в нормотонусе, сокращений и схваток нет.
4. Периодические изменения сердечного ритма
Акцелерации – это «пики» на графике, вершина которых обращена вверх, это означает учащение сердцебиения ребенка. Акцелерации соотносят с графиком шевелений (шевеление в норме вызывает учащение ЧСС) и с гистерограммой (повышение тонуса матки или схватки вызывают учащение ЧСС). Акцелераций на нормальной КТГ должно быть не менее двух за 40 минут.
Децелерации – это «пики» с вершиной вниз или «ямы» на тахограмме:
Ранние или I тип возникают вместе со схваткой или через несколько секунд, имеют плавные границы, они могут быть одиночными и неглубокими, но не должны идти группами. Это один из признаков сдавления пуповины при схватке.
Это один из признаков сдавления пуповины при схватке.
Поздние или II тип регистрируются после схватки, глубокие и их продолжительность больше, чем время схватки (вычисляются в секундах по оси абсцисс), в норме не должны быть, это признак нарушения кровообращения в плаценте.
Вариабельные или III тип имеют различную форму, нет синхронизации со схватками. Могут свидетельствовать о компрессии пуповины, маловодии, единичные могут появляться во время шевелений.
Для оценки КТГ в баллах существует таблица. Этот метод называется оценкой по Фишеру.
| Параметры ЧСС | Баллы | ||
| 0 | 1 | 2 | |
| Базальная ЧСС | >180 | 100-120 160-180 | 120-160 |
| Вариабельность (частота осцилляций в минуту) | Менее 3 | 3-6 | 6 и более |
| Амплитуда осцилляций | 5 и менее | 5-8 или более 25 | 9-25 |
| Изменения ЧСС: Акцелерации Децелерации | Отсутствуют Поздние, длительные или вариабельные | Периодические Поздние, кратковременные или вариабельные | Спорадические Ранние или отсутствуют |
8-10 баллов нормальная КТГ
6-7 сомнительный тип
Менее 6 – патологический тип КТГ.
Тактика
При нормотипе КТГ беременность или роды продолжают вести в соответствии со стандартами. Это означает, что ребенок не страдает, и беременность/роды протекают благополучно.
При сомнительном типе КТГ оценивают суммарный риск, то есть состояние матери, предшествующие заболевания и осложнения беременности (преэклампсия, рубец на матке после кесарева сечения). В зависимости от этого принимают решение. Здесь стоит выбор между дальнейшим динамическим наблюдением и срочным родоразрешением. Для решения этой задачи проводят дополнительные обследования (УЗИ-контроль с допплерометрией, осмотр консилиумом врачей).
Патологический тип КТГ
свидетельствует о выраженной гипоксии плода и является показанием для экстренного родоразрешения. Если это состояние зафиксировано при беременности или с началом родов, то родоразрешение производится путем операции кесарева сечения. Если гипоксия возникает в результате слабости потужного периода (непосредственно рождение ребенка), то накладываются акушерские щипцы или производится вакуум-экстракция плода.
Нам повезло, что сейчас существует безвредный и информативный метод исследования, который можно использовать так часто, как только понадобится. Выполняйте рекомендации врача, сообщайте о своих жалобах и сомнениях и не ленитесь прийти на контроль КТГ, если вам назначено. Мы желаем вам благополучной беременности и легких родов. Следите за собой и будьте здоровы!
Врач акушер-гинеколог Петрова А.В.
Транскрипция и ядерный транспорт тринуклеотидных повторов CAG / CTG в дрожжах
Nucleic Acids Res. 2002 Aug 15; 30 (16): 3540–3547.
Unité de Génétique Moléculaire des Levures (URA 2171 CNRS и UFR 927 Université Pierre et Marie Curie) Institut Pasteur, 25 rue du Dr Roux, 75724 Paris cedex 15, France
a Кому следует обращаться. Тел .: +33 1 40 61 34 54; Факс: +33 1 40 61 34 56; Электронная почта: rf.ruetsap@erbafeПоступила 21 мая 2002 г .; Пересмотрено 26 июня 2002 г .; Принята в печать 26 июня 2002 г.
Эта статья цитируется в других статьях в PMC.
Abstract
Тринуклеотидные повторы участвуют в нескольких неврологических расстройствах у человека. Последовательности ДНК, содержащие повторы CAG / CTG, склонны к проскальзыванию во время репликации и репарации двухцепочечных разрывов. Эффекты тринуклеотидных повторов на транскрипцию и ядерный экспорт были проанализированы in vivo в дрожжах. Транскрипция тринуклеотидного повтора CAG / CTG в 3′-нетранслируемой области репортерного гена URA3 приводит к транскрипции информационных РНК на несколько килобаз длиннее ожидаемого.Эти длинные мРНК образуются легче, когда транскрибируются повторы CAG, а не CTG. Транскрипты, содержащие CAG или CUG, обнаруживают негомогенную клеточную локализацию. Мы предполагаем, что длинные мРНК возникают в результате проскальзывания транскрипции, и обсуждаем возможные последствия для болезней человека.
ВВЕДЕНИЕ
Среди неврологических расстройств человека, связанных с разрастанием тринуклеотидных повторов, наиболее частым триплетом, который выявляется, является CAG и его комплементарная последовательность CTG (1). Повторы CAG почти всегда обнаруживаются в открытых рамках считывания (2), тогда как расширения CTG расположены в 3′-нетранслируемой области (3′-UTR) гена DMPK при миотонической дистрофии 1 типа (3,4) и в SCA8 (5). Транскрипция расширенного аллеля DMPK у человека существенно не снижается. Тем не менее, было обнаружено, что экспансии CUG нарушают процессинг пре-мРНК (6) и что CUG-содержащие РНК накапливаются в ядре в виде фокусов (7-9). Миотоническая дистрофия 2, вызванная экспансией тетрануклеотидного повтора CCTG, также обнаруживает ядерную задержку транскриптов (10).Поскольку белки, принадлежащие к семейству, участвующему в мышечной дифференцировке, специфически связывают РНК, содержащие повторы CUG, было высказано предположение, что аномальное рекрутирование этих белков может быть ответственным за патологию DM-1 (11). Удерживание в ядре специфично для повторов CUG, поскольку мРНК, содержащие повторы CAG, при других нарушениях тринуклеотидных повторов, по-видимому, правильно экспортируются.
Повторы CAG почти всегда обнаруживаются в открытых рамках считывания (2), тогда как расширения CTG расположены в 3′-нетранслируемой области (3′-UTR) гена DMPK при миотонической дистрофии 1 типа (3,4) и в SCA8 (5). Транскрипция расширенного аллеля DMPK у человека существенно не снижается. Тем не менее, было обнаружено, что экспансии CUG нарушают процессинг пре-мРНК (6) и что CUG-содержащие РНК накапливаются в ядре в виде фокусов (7-9). Миотоническая дистрофия 2, вызванная экспансией тетрануклеотидного повтора CCTG, также обнаруживает ядерную задержку транскриптов (10).Поскольку белки, принадлежащие к семейству, участвующему в мышечной дифференцировке, специфически связывают РНК, содержащие повторы CUG, было высказано предположение, что аномальное рекрутирование этих белков может быть ответственным за патологию DM-1 (11). Удерживание в ядре специфично для повторов CUG, поскольку мРНК, содержащие повторы CAG, при других нарушениях тринуклеотидных повторов, по-видимому, правильно экспортируются.
CTG-повторов вызывают временные паузы в элонгации РНК in vitro (12). Однако неизвестно, возникают ли аналогичные паузы in vivo, , и могут ли они способствовать проскальзыванию транскрипта во время элонгации.Проскальзывание транскрипции наблюдалось in vitro (13–16), in vivo у Escherichia coli (17,18) и у пациентов с болезнью Альцгеймера и Дауна (19,20), но никогда не сообщалось для тринуклеотидных повторов.
Мы использовали дрожжи Saccharomyces cerevisiae для исследования эффектов длинных повторов CAG или CTG на транскрипцию и на ядерно-цитоплазматический транспорт репортерной мРНК, содержащей тринуклеотидные повторы в ее 3′-UTR. Мы обнаружили, что транскрипты, содержащие CAG и CUG, длиннее ожидаемого.Формирование этих длинных транскриптов зависит от последовательности и длины. Кроме того, мы показываем, что эти мРНК неравномерно распределены в клетке. Мы обсуждаем возможные последствия этих двух наблюдений для патологий человека, связанных с повторами CAG / CTG.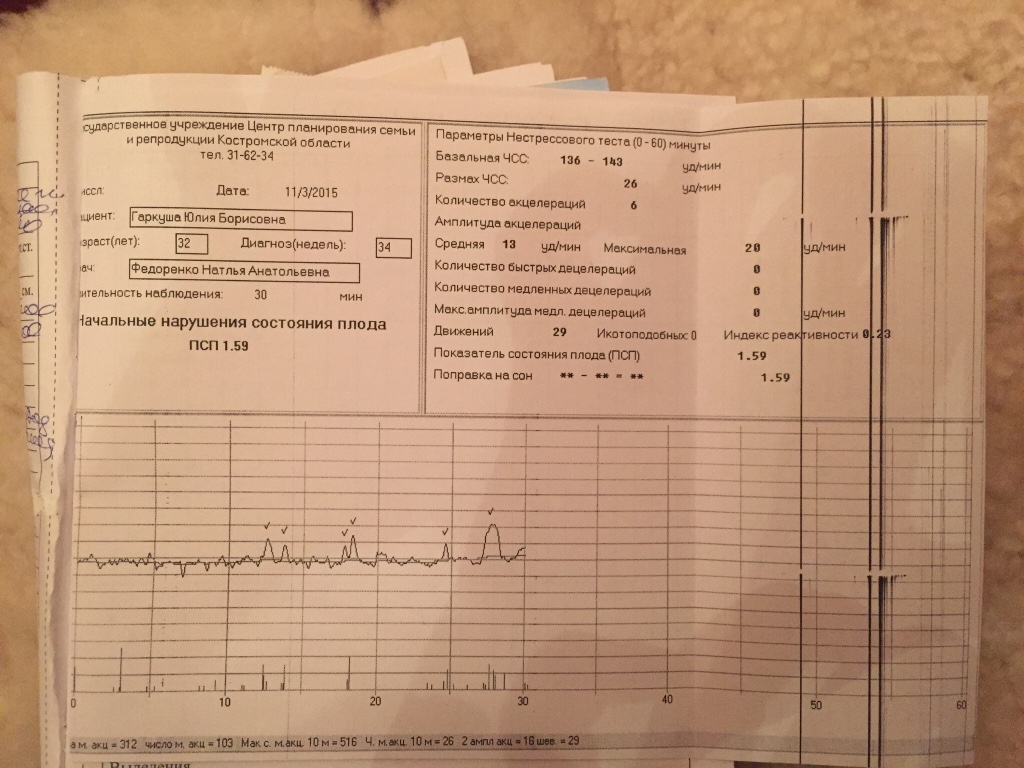
МАТЕРИАЛЫ И МЕТОДЫ
Плазмиды и штаммы
Плазмида pCMha191 была получена из pCM184 (21) путем замены CEN-ARS-фрагмента на 2 мкм репликационный ориджин, созданного с помощью ПЦР, и путем клонирования олигонуклеотида, содержащего метку HA13, расположенную ниже по течению. Промотор CYC1 (J.Бойе, неопубликованные результаты). Ген URA3 амплифицировали с помощью ПЦР с использованием ДНК-полимеразы Pfu (Stratagene) и клонировали в рамке с меткой HA в pCMha191 с получением плазмиды pE186. pTRI625 был сконструирован путем клонирования участка λ ДНК из Pst I размером 300 п.н. в сайт Pst I, расположенный в 3′-UTR URA3 pE186 (фиг. A). pTRI626 был сконструирован путем клонирования участка λ ДНК Pst I размером 700 п.н. в тот же сайт. pTRI627 был сконструирован путем клонирования фрагмента ДНК Bam HI– Xba I размером 300 п.н., содержащего (CAG) 98 тринуклеотидный повтор из pTRI620 (J.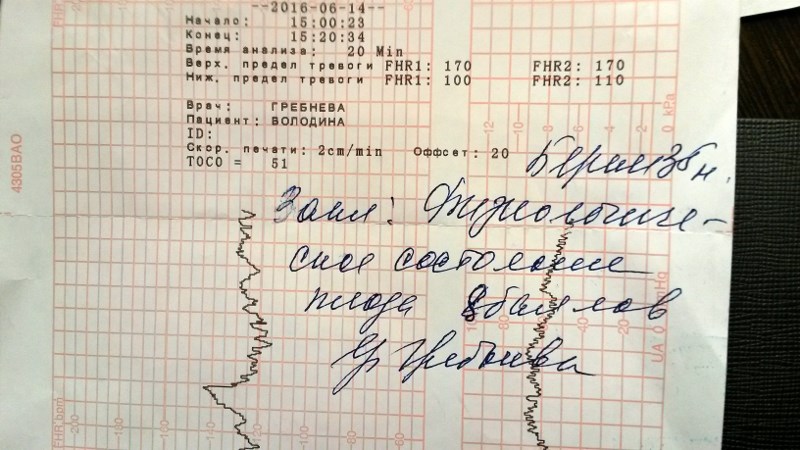 Boyer и B.Dujon, неопубликованные данные) между сайтами Bgl II и Nhe I, расположенными в 3′-UTR URA3 pE186. pTRI628 был сконструирован путем клонирования фрагмента ДНК Bam HI– Pst I размером 300 п.н., содержащего (CTG) 98 тринуклеотидный повтор из pRW3216 (22) между сайтами Bgl II и Pst I, расположенными в URA3 3′-UTR из pE186. pTRI629 был сконструирован путем клонирования фрагмента ДНК длиной 780 п.н., содержащего (CTG) 255 тринуклеотидный повтор из pRW3222 (22) между теми же сайтами в той же плазмиде.pTRI637 был сконструирован путем клонирования тринуклеотидного повтора размером 300 п.н. (TTG) из pTRI622 (23) между сайтами Pst I и Nhe I, расположенными в 3′-UTR URA3 pE186. Трансформации плазмид pE186 и pTRI625–637 проводили в штамме FYBL1-22B ( MAT a ura3 Δ851 trp1 Δ63 his3 Δ200 lys2 Δ202) (24), используя одностадийный метод (25 ).
Boyer и B.Dujon, неопубликованные данные) между сайтами Bgl II и Nhe I, расположенными в 3′-UTR URA3 pE186. pTRI628 был сконструирован путем клонирования фрагмента ДНК Bam HI– Pst I размером 300 п.н., содержащего (CTG) 98 тринуклеотидный повтор из pRW3216 (22) между сайтами Bgl II и Pst I, расположенными в URA3 3′-UTR из pE186. pTRI629 был сконструирован путем клонирования фрагмента ДНК длиной 780 п.н., содержащего (CTG) 255 тринуклеотидный повтор из pRW3222 (22) между теми же сайтами в той же плазмиде.pTRI637 был сконструирован путем клонирования тринуклеотидного повтора размером 300 п.н. (TTG) из pTRI622 (23) между сайтами Pst I и Nhe I, расположенными в 3′-UTR URA3 pE186. Трансформации плазмид pE186 и pTRI625–637 проводили в штамме FYBL1-22B ( MAT a ura3 Δ851 trp1 Δ63 his3 Δ200 lys2 Δ202) (24), используя одностадийный метод (25 ).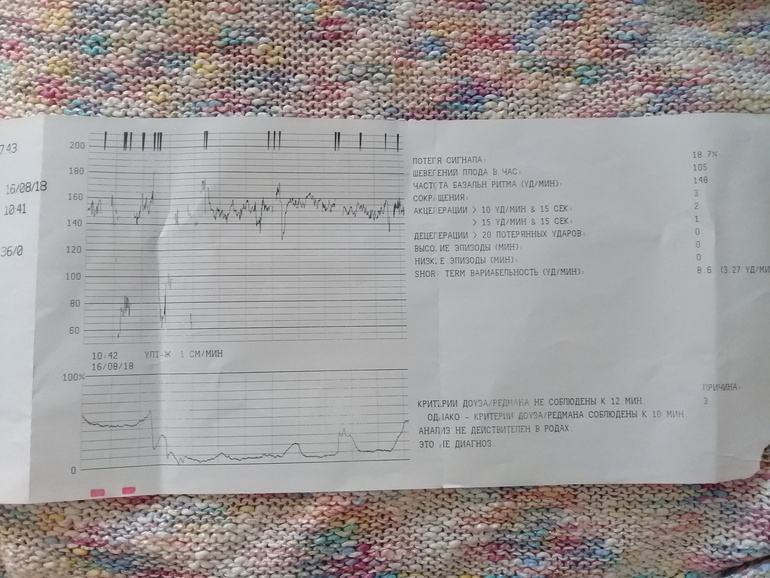 Плазмиды pE186, pTRI627 и pTRI628 были также трансформированы в штамм FYEF95 ( MAT a ura3 Δ851 trp1 Δ63 leu2 Δ 1 nup145 Δ nup145N-LEU2 ) с использованием того же протокола (26).pTRI644 и pTRI645 были соответственно сконструированы путем клонирования фрагмента Eco RI– Bgl II из pTRI627 или pTRI628 в плазмиду pTRI643, содержащую дрожжевой ген ARG2 , фланкированный двумя сайтами Not I, чтобы обеспечить линеаризацию перед трансформацией дрожжи. Плазмиды pTRI644 и pTRI645 трансформировали в штамм FYBL1-22B после линеаризации Not I, чтобы получить штаммы GFR108 и GFR109, соответственно, с использованием протокола, разработанного Gietz et al .(27).
Плазмиды pE186, pTRI627 и pTRI628 были также трансформированы в штамм FYEF95 ( MAT a ura3 Δ851 trp1 Δ63 leu2 Δ 1 nup145 Δ nup145N-LEU2 ) с использованием того же протокола (26).pTRI644 и pTRI645 были соответственно сконструированы путем клонирования фрагмента Eco RI– Bgl II из pTRI627 или pTRI628 в плазмиду pTRI643, содержащую дрожжевой ген ARG2 , фланкированный двумя сайтами Not I, чтобы обеспечить линеаризацию перед трансформацией дрожжи. Плазмиды pTRI644 и pTRI645 трансформировали в штамм FYBL1-22B после линеаризации Not I, чтобы получить штаммы GFR108 и GFR109, соответственно, с использованием протокола, разработанного Gietz et al .(27).
Экспериментальные системы, использованные в данной работе. ДНК ( A ) λ или тринуклеотидного повтора клонировали в 3′-UTR гена URA3 под контролем регулируемого промотора tetOn / CYC1 . Тринуклеотидные повторы или контрольные последовательности клонировали между стоп-кодоном и терминатором транскрипции CYC1 . ( B ) Плазмиды pTRI627 и pTRI628, показанные на (A), удаленные из-за их ориджина репликации, были интегрированы в локус ARG2 (пунктирная рамка).
Тринуклеотидные повторы или контрольные последовательности клонировали между стоп-кодоном и терминатором транскрипции CYC1 . ( B ) Плазмиды pTRI627 и pTRI628, показанные на (A), удаленные из-за их ориджина репликации, были интегрированы в локус ARG2 (пунктирная рамка).
Экстракция тотальной РНК и Нозерн-блоттинг
Используемая процедура описана Ричардом и др. . (28). Образцы РНК денатурируют в 80% формамиде в течение 5 минут при 95 ° C перед нанесением на денатурирующий агарозный гель, содержащий 0,6% формальдегида. Зонды были случайным образом помечены с использованием [α- 32 P] dATP для зонда URA3 и [α- 32 P] dCTP для зонда CAG / CTG. Все количественные оценки были выполнены с использованием Phosphorimager (Molecular Dynamics) и программного обеспечения ImageQuant 5.1.
Получение полиаденилированных РНК
Олиго (dT) -целлюлозу типа 7 (Pharmacia) промывали 0,1 М NaOH в течение 30 мин и уравновешивали в буфере «H» (0,5 М NaCl, 10 мМ Трис, pH 7,5, 2 мМ ЭДТА, 0,1% SDS).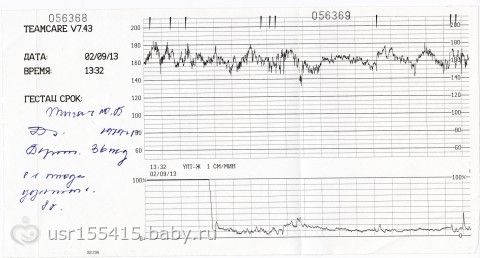 Аликвоту общей РНК в 500 мкг денатурировали в течение 10 минут при 65 ° C, ресуспендировали в 300 мкл буфера «H» и инкубировали в присутствии 125 мг олиго (dT) -целлюлозы в течение 30 минут при комнатной температуре с осторожным трясется. Три промывки (по 5 минут каждая при комнатной температуре) были выполнены в буфере «M» (0.15 M NaCl, 10 мМ Трис pH 7,5, 2 мМ EDTA и 0,1% SDS). МРНК поли (A) + элюировали в буфере «L» (2 мМ EDTA и 0,1% SDS) при 65 ° C в течение 5 минут. Аликвоту 1 мкг поли (A) + РНК загружали в гели для последующего нозерн-блоттинга.
Аликвоту общей РНК в 500 мкг денатурировали в течение 10 минут при 65 ° C, ресуспендировали в 300 мкл буфера «H» и инкубировали в присутствии 125 мг олиго (dT) -целлюлозы в течение 30 минут при комнатной температуре с осторожным трясется. Три промывки (по 5 минут каждая при комнатной температуре) были выполнены в буфере «M» (0.15 M NaCl, 10 мМ Трис pH 7,5, 2 мМ EDTA и 0,1% SDS). МРНК поли (A) + элюировали в буфере «L» (2 мМ EDTA и 0,1% SDS) при 65 ° C в течение 5 минут. Аликвоту 1 мкг поли (A) + РНК загружали в гели для последующего нозерн-блоттинга.
Обработка РНКазой и ДНКазой
Для обработки РНКазой 5 мкг общей РНК инкубировали в 40 мМ Трис-HCl, pH 7,5, 6 мМ MgCl 2 , с 0,25 мкг РНКазы A, свободной от ДНКазы (Boehringer), в конечном объем 6 мкл, в течение 20 мин при 37 ° C.В качестве альтернативы, 5 мкг общей РНК инкубировали в 800 мМ NaOH (конечный объем 6 мкл) в течение 20 минут при 65 ° C для щелочного гидролиза. Нейтрализация достигалась добавлением 5 мкл 1 М трис-HCl, pH 7,5, и 3 мкл 2 н. HCl. Обработку ДНКазой проводили на 5 мкг общей РНК, инкубированной в 40 мМ Трис-HCl, pH 7,5, 6 мМ MgCl 2 , с 5 ед. FPLC-чистой ДНКазы I (Pharmacia) и 5 ед. Ингибитора РНКазы (Promega) в конечном итоге. объем 6,5 мкл в течение 20 мин при 37 ° C.
HCl. Обработку ДНКазой проводили на 5 мкг общей РНК, инкубированной в 40 мМ Трис-HCl, pH 7,5, 6 мМ MgCl 2 , с 5 ед. FPLC-чистой ДНКазы I (Pharmacia) и 5 ед. Ингибитора РНКазы (Promega) в конечном итоге. объем 6,5 мкл в течение 20 мин при 37 ° C.
Флуоресценция
in situ гибридизация (FISH)мРНК CUG и CAG были обнаружены, как описано Jensen et al .(29). Каждый образец клеток гибридизовали с 0,5 пмоль Cy3-меченными праймерами CAG или CTG 21mer. Ядерную ДНК метили 0,1 мкг / мл DAPI. Зонд Cy3-dT (30-мерный) использовали для нормализации соотношения ядерных сигналов. Были количественно определены сигналы зондов CAG, CTG и oligo-dT (ImageQuant 5.1). Ядерные сигналы, обнаруженные каждым зондом, количественно оценивали с помощью окрашивания DAPI. Отношения ядерный сигнал / общий сигнал существенно не различались между зондами для каждого штамма.
РЕЗУЛЬТАТЫ
Тринуклеотидные повторы CAG и CTG генерируют длинные РНК
Мы разработали экспериментальную систему на дрожжах, в которой различные повторы были клонированы в 3′-UTR гена URA3 (рис. ). Несколько эписомальных конструкций, содержащих CAG 98 , CTG 98 , CTG 255 и TTG 87 , а также участки ДНК λ длиной 300 или 700 п.н. в качестве контроля трансформировали в дрожжи. Суммарные РНК от трансформантных штаммов исследовали с помощью нозерн-блоттинга (фиг. A). Для каждого штамма был обнаружен транскрипт URA3 ожидаемого размера. Удивительно, но мазки, превышающие ожидаемые размеры транскриптов на несколько тысяч пар оснований, также были обнаружены у трех штаммов, содержащих повторы CAG или CUG (рис.А). Все виды РНК, обнаруженные в общей РНК, также были обнаружены во фракции поли (A) + (рис. B). Было обнаружено, что как зрелые РНК размера, так и мазки с высокой молекулярной массой гибридизуются с зондом CAG / CTG, что указывает на то, что соответствующие молекулы РНК содержат повторы CAG или CUG (рис. C). Гибридизация с ДНК-зондом, покрывающим область сразу после сайта терминации транскрипции, не показала детектируемого сигнала (данные не показаны), что указывает на то, что мазки, содержащие CAG или CUG, не выходят за пределы нормального сайта терминации.
). Несколько эписомальных конструкций, содержащих CAG 98 , CTG 98 , CTG 255 и TTG 87 , а также участки ДНК λ длиной 300 или 700 п.н. в качестве контроля трансформировали в дрожжи. Суммарные РНК от трансформантных штаммов исследовали с помощью нозерн-блоттинга (фиг. A). Для каждого штамма был обнаружен транскрипт URA3 ожидаемого размера. Удивительно, но мазки, превышающие ожидаемые размеры транскриптов на несколько тысяч пар оснований, также были обнаружены у трех штаммов, содержащих повторы CAG или CUG (рис.А). Все виды РНК, обнаруженные в общей РНК, также были обнаружены во фракции поли (A) + (рис. B). Было обнаружено, что как зрелые РНК размера, так и мазки с высокой молекулярной массой гибридизуются с зондом CAG / CTG, что указывает на то, что соответствующие молекулы РНК содержат повторы CAG или CUG (рис. C). Гибридизация с ДНК-зондом, покрывающим область сразу после сайта терминации транскрипции, не показала детектируемого сигнала (данные не показаны), что указывает на то, что мазки, содержащие CAG или CUG, не выходят за пределы нормального сайта терминации. Полосы, соответствующие мРНК нормального размера и мазкам, не были обнаружены после обработки РНКазой или щелочным гидролизом, но все еще были видны после обработки ДНКазой I, подтверждая, что мазки не были связаны с загрязнением ДНК (см. Материалы и методы). Молекулы РНК, обнаруженные как мазки с высокой молекулярной массой, не транскрибировались из участков увеличенных повторов ДНК CAG / CTG, поскольку не было обнаружено увеличения числа повторов CAG / CTG в различных штаммах, проанализированных с помощью саузерн-блоттинга (дополнительный материал).Мазки, видимые ниже основной полосы при ожидаемом размере 1,4 т.п.н., связаны либо с деградацией мРНК (как это обычно наблюдается с обильными транскриптами) (30), либо, в качестве альтернативы, с повторением сокращений тракта на уровне ДНК (дополнительный материал). Из этой серии экспериментов мы пришли к выводу, что длинные РНК, обнаруженные в виде мазков, соответствуют поли (A) + РНК, транскрибируемым с матрицы ДНК нормального размера, содержащей повторы CAG или CUG.
Полосы, соответствующие мРНК нормального размера и мазкам, не были обнаружены после обработки РНКазой или щелочным гидролизом, но все еще были видны после обработки ДНКазой I, подтверждая, что мазки не были связаны с загрязнением ДНК (см. Материалы и методы). Молекулы РНК, обнаруженные как мазки с высокой молекулярной массой, не транскрибировались из участков увеличенных повторов ДНК CAG / CTG, поскольку не было обнаружено увеличения числа повторов CAG / CTG в различных штаммах, проанализированных с помощью саузерн-блоттинга (дополнительный материал).Мазки, видимые ниже основной полосы при ожидаемом размере 1,4 т.п.н., связаны либо с деградацией мРНК (как это обычно наблюдается с обильными транскриптами) (30), либо, в качестве альтернативы, с повторением сокращений тракта на уровне ДНК (дополнительный материал). Из этой серии экспериментов мы пришли к выводу, что длинные РНК, обнаруженные в виде мазков, соответствуют поли (A) + РНК, транскрибируемым с матрицы ДНК нормального размера, содержащей повторы CAG или CUG.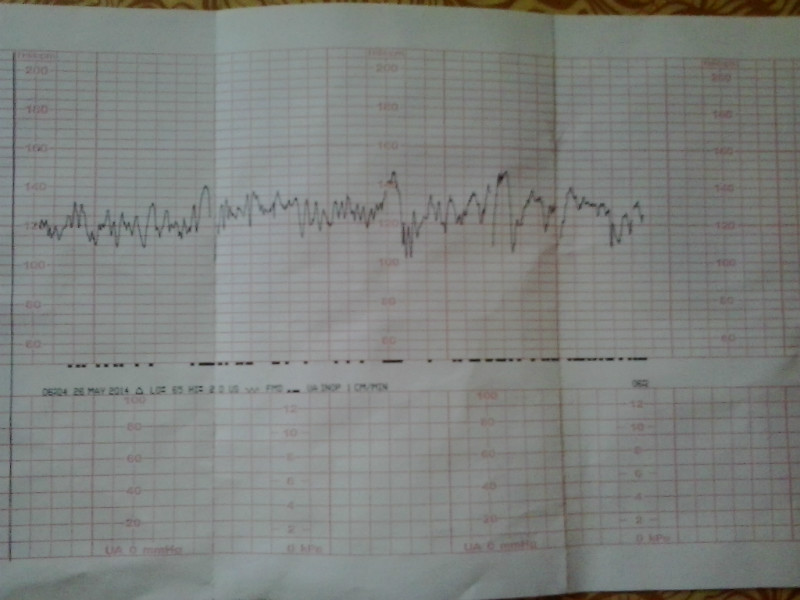 В дальнейшем они будут называться L-РНК.
В дальнейшем они будут называться L-РНК.
(напротив) Нозерн-блоты, показывающие зрелые мРНК URA3 и L-РНК. ( A ) Суммарные РНК штаммов дрожжей, содержащие конструкции, показанные на фигуре A, гибридизированные с зондом URA3 . Маркеры размера РНК указаны слева. Транскрипт CUG 98 длиннее транскрипта CAG 98 из-за дополнительной части полилинкера pBluescript. В контрольном штамме FYBL1-2B / pTRI626 более низкий сигнал, возможно, соответствует альтернативному сайту терминации или процессинга, расположенному в 700 п.н. ДНК λ.L-РНК видны в виде мазков, простирающихся от зрелого транскрипта URA3 (~ 1,4 т.п.н.) до> 4,4 т.п.н. ( B ) мРНК Poly (A) + , очищенные из тех же штаммов и гибридизованные с тем же зондом. Нижняя полоса в штамме FYBL1-2B / pTRI626 не обнаруживается во фракции поли (A) + . ( C ) Тот же блот, что и в (B), очищенный и повторно гибридизированный с зондом CAG / CTG. ( D ) Количественная оценка радиоактивности для общих РНК, гибридизованных с зондом URA3 .Темно-серая область соответствует молекулам мРНК URA3 и ожидаемого размера. Светло-серая область соответствует L-РНК. Мазки меньшего размера, чем ожидалось, соответствуют деградации РНК. Локальное уменьшение сигнала соответствует конкуренции со стороны большого количества молекул рРНК. ( E ) Нозерн-блоттинг, сравнивающий образцы РНК, выделенные из штаммов, содержащих либо эписомальную конструкцию URA3 -UUG 87 , хромосомную конструкцию URA3 -CAG 98 или хромосомную конструкцию URA3 -CUG 98 .Транскрипт UUG 87 длиннее транскрипта CUG 98 из-за дополнительных участков ДНК размером 150 п.н., происходящих из исходного вектора, содержащего участок повтора TTG 87 . В этом конкретном эксперименте количества L-РНК составляли, соответственно, 16% (UUG 87 ), 18% (CUG 98 ) и 85% (CAG 98 ).
( D ) Количественная оценка радиоактивности для общих РНК, гибридизованных с зондом URA3 .Темно-серая область соответствует молекулам мРНК URA3 и ожидаемого размера. Светло-серая область соответствует L-РНК. Мазки меньшего размера, чем ожидалось, соответствуют деградации РНК. Локальное уменьшение сигнала соответствует конкуренции со стороны большого количества молекул рРНК. ( E ) Нозерн-блоттинг, сравнивающий образцы РНК, выделенные из штаммов, содержащих либо эписомальную конструкцию URA3 -UUG 87 , хромосомную конструкцию URA3 -CAG 98 или хромосомную конструкцию URA3 -CUG 98 .Транскрипт UUG 87 длиннее транскрипта CUG 98 из-за дополнительных участков ДНК размером 150 п.н., происходящих из исходного вектора, содержащего участок повтора TTG 87 . В этом конкретном эксперименте количества L-РНК составляли, соответственно, 16% (UUG 87 ), 18% (CUG 98 ) и 85% (CAG 98 ).
Количество L-РНК определяли количественно (рис. D). Для штаммов, содержащих повторы CAG 98 или CUG 98 , 40–50% всех мРНК URA3 были обнаружены как L-РНК и до 69% для штамма, содержащего повтор CUG 255 (рис.А).
( A ) Относительное количество молекул L-РНК по сравнению с общим количеством транскриптов URA3 , обнаруженных в каждом штамме с помощью зонда URA3 . Количественная оценка была проведена, как показано на рисунке D. Отношения рассчитываются как [светло-серая область / общие (светлые + темные) серые области] (в%). ( B ) Соотношения сигналов L-РНК, детектируемых зондом CAG / CTG, по сравнению с такими же сигналами, детектируемыми зондом URA3 . Ожидаемый размер мРНК URA3 использовался в качестве внутреннего стандарта, и отношение сигнала URA3 , обнаруженного с помощью зонда CAG / CTG, к такому же сигналу, обнаруженному с помощью зонда URA3 , было произвольно установлено на 1 (показано белым: УРА3 ).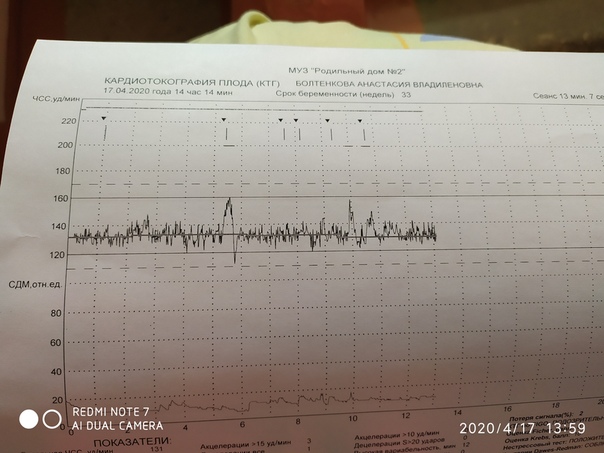 Соотношения для конструкций CAG и CUG показаны светло-серым и темно-серым цветом соответственно. Кинетика гибридизации на мембране в присутствии избытка зонда (примерно 10 8 молекул РНК на мембране по сравнению с 4,5 × 10 11 молекул зонда) зависит от концентрации зонда (40). Поскольку использовались одинаковые концентрации зонда, кинетика гибридизации схожа, и результирующие сигналы пропорциональны только количеству молекул РНК на мембране. Каждая молекула CAG / CTG также несет URA3 , но соотношение CAG или CTG к URA3 выше 1.Это означает, что с РНК связано больше молекул зондов CAG / CTG, чем молекул зондов URA3 , поэтому часть CAG / CTG длиннее, чем часть URA3 . Обратите внимание, что более высокое соотношение с CAG 98 предполагает, что CAG-содержащие L-РНК длиннее, чем те, которые продуцируются в CUG-содержащих штаммах. Умеренное увеличение соотношения CUG 255 по сравнению с CUG 98 связано с более высоким сигналом РНК зрелого размера CUG 255 с зондом CAG / CTG (сравните сигнал зрелого размера URA3 РНК в полосе URA3 -CUG 255 на рис.
Соотношения для конструкций CAG и CUG показаны светло-серым и темно-серым цветом соответственно. Кинетика гибридизации на мембране в присутствии избытка зонда (примерно 10 8 молекул РНК на мембране по сравнению с 4,5 × 10 11 молекул зонда) зависит от концентрации зонда (40). Поскольку использовались одинаковые концентрации зонда, кинетика гибридизации схожа, и результирующие сигналы пропорциональны только количеству молекул РНК на мембране. Каждая молекула CAG / CTG также несет URA3 , но соотношение CAG или CTG к URA3 выше 1.Это означает, что с РНК связано больше молекул зондов CAG / CTG, чем молекул зондов URA3 , поэтому часть CAG / CTG длиннее, чем часть URA3 . Обратите внимание, что более высокое соотношение с CAG 98 предполагает, что CAG-содержащие L-РНК длиннее, чем те, которые продуцируются в CUG-содержащих штаммах. Умеренное увеличение соотношения CUG 255 по сравнению с CUG 98 связано с более высоким сигналом РНК зрелого размера CUG 255 с зондом CAG / CTG (сравните сигнал зрелого размера URA3 РНК в полосе URA3 -CUG 255 на рис.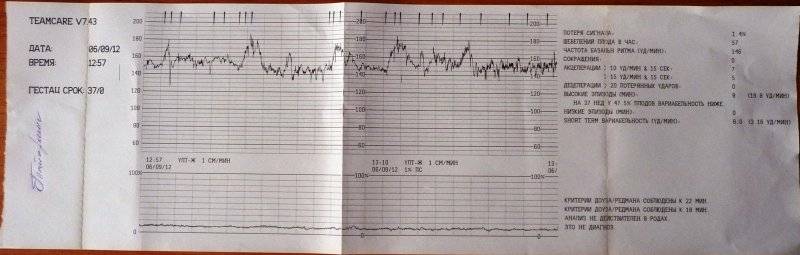 C и D).
C и D).
Чтобы определить, играет ли природа повторяющейся последовательности роль в процессе генерации L-РНК, мы экспрессировали повтор TTG 87 в том же положении (рис. A). В отличие от повторов CAG и CTG, повторы TTG n (или их дополнительные повторы CAA n ) не образуют стабильных вторичных структур in vitro . В штаммах, содержащих конструкцию URA3 -TTG 87 , уровень L-РНК был снижен до 19%, что значительно ниже уровней, наблюдаемых с URA3 -CAG 98 или URA3 -CTG 98 конструкции (рис. E и A).Таким образом, мы пришли к выводу, что природа повторяющейся последовательности влияет на уровень L-РНК и что, скорее всего, in vivo стабильные вторичные структуры ДНК способствуют образованию L-РНК.
Чтобы определить, влияет ли экспрессия из хромосомного местоположения на образование L-РНК по сравнению с многокопийной плазмидой, в дрожжах был интегрирован ген URA3 , содержащий CAG 98 или CTG 98 повторов в его 3′-UTR. хромосома X в локусе ARG2 (рис.Б). Общее количество транскриптов URA3 , детектированных с помощью этой конструкции, было ниже, чем с помощью эписомальных конструкций, однако поразительное снижение количества L-РНК наблюдалось при продуцировании мРНК URA3 -CUG 98 (штамм GFR109) по сравнению с с мРНК URA3 -CAG 98 (штамм GFR108) (рис.). Разница в относительном количестве L-РНК между двумя хромосомными конструкциями более выражена, чем с эписомными конструкциями. Мы пришли к выводу, что ориентация повторяющейся последовательности по сравнению с направлением транскрипции также важна для генерации L-РНК.
хромосома X в локусе ARG2 (рис.Б). Общее количество транскриптов URA3 , детектированных с помощью этой конструкции, было ниже, чем с помощью эписомальных конструкций, однако поразительное снижение количества L-РНК наблюдалось при продуцировании мРНК URA3 -CUG 98 (штамм GFR109) по сравнению с с мРНК URA3 -CAG 98 (штамм GFR108) (рис.). Разница в относительном количестве L-РНК между двумя хромосомными конструкциями более выражена, чем с эписомными конструкциями. Мы пришли к выводу, что ориентация повторяющейся последовательности по сравнению с направлением транскрипции также важна для генерации L-РНК.
Нозерн-блоттинг тотальных РНК, экстрагированных из штаммов GFR108 и GFR109, экспрессирующих хромосомные конструкции URA3 -CAG 98 или URA3 -CUG 98 , соответственно, гибридизованных с URA3 или ACT1 зонд. Сигнал актина использовали для сравнения общего количества мРНК в обеих дорожках (см. Текст). Количественная оценка L-РНК выполняется, как описано на рисунке A. Дискретные полосы соответствуют конкуренции большого количества молекул рРНК, что приводит к локальному снижению сигнала.
Текст). Количественная оценка L-РНК выполняется, как описано на рисунке A. Дискретные полосы соответствуют конкуренции большого количества молекул рРНК, что приводит к локальному снижению сигнала.
Поскольку повторы CAG / CTG имеют склонность к образованию стабильных вторичных структур in vitro (31–34), а повторы CUG также могут образовывать аналогичные стабильные вторичные структуры (35), мы не могли исключить, что L-РНК представляют собой структурированные РНК. которые аберрантно мигрируют в агарозных гелях. Однако штамм GFR108 ( URA3 -CAG 98 ) демонстрирует гораздо более высокую долю L-РНК, чем штамм GFR109 ( URA3 -CUG 98 ). Поскольку шпильки, образованные повторами CUG, более стабильны, чем шпильки, образованные повторами CAG (35), мы могли бы ожидать противоположного результата, если бы L-РНК действительно были вторичными структурами.Кроме того, мы обнаружили, что сигнал L-РНК, обнаруженный с помощью зонда CAG / CTG, выше, чем тот же сигнал, обнаруженный с помощью зонда URA3 (рис. B), с сопоставимой удельной активностью (зонд CAG / CTG, 9,2 × 10 7 имп / мин / мкг; URA3 зонд, 7,7 × 10 8 имп / мин / мкг). Это различие более важно между сигналами CAG и URA3 , чем между сигналами CUG и URA3 , предполагая, что молекулы, содержащие CAG, действительно более длинные молекулы, чем молекулы, содержащие CUG.Это видно на рисунках A, B и E (на рисунке C показан тот же гель, что и на рисунке B, повторно гибридизированный с зондом CAG / CTG), где большинство молекул L-РНК CAG имеют более высокий молекулярный вес, чем L-РНК CUG. молекулы. Измерения были выполнены в двух независимых экспериментах, и расчет стандартной ошибки показывает, что разница между двумя отношениями статистически значима (рис. B). В попытке клонировать и секвенировать эти длинные молекулы РНК, нам не удалось осуществить обратную транскрипцию и амплифицировать полноразмерную кДНК (дополнительный материал).
B), с сопоставимой удельной активностью (зонд CAG / CTG, 9,2 × 10 7 имп / мин / мкг; URA3 зонд, 7,7 × 10 8 имп / мин / мкг). Это различие более важно между сигналами CAG и URA3 , чем между сигналами CUG и URA3 , предполагая, что молекулы, содержащие CAG, действительно более длинные молекулы, чем молекулы, содержащие CUG.Это видно на рисунках A, B и E (на рисунке C показан тот же гель, что и на рисунке B, повторно гибридизированный с зондом CAG / CTG), где большинство молекул L-РНК CAG имеют более высокий молекулярный вес, чем L-РНК CUG. молекулы. Измерения были выполнены в двух независимых экспериментах, и расчет стандартной ошибки показывает, что разница между двумя отношениями статистически значима (рис. B). В попытке клонировать и секвенировать эти длинные молекулы РНК, нам не удалось осуществить обратную транскрипцию и амплифицировать полноразмерную кДНК (дополнительный материал).
CAG и CTG-повторы, экспортируются, но образуют кластеры в дрожжевых клетках
Чтобы определить, экспортировались ли мРНК, содержащие тринуклеотидный повтор, в цитоплазму, мы проанализировали локализацию URA3 мРНК с помощью FISH с использованием Cy3-меченного CAG или олигонуклеотиды CTG. Способность зонда проникать в ядро оценивали на штамме, который, как известно, нарушает ядерный экспорт мРНК (Δ nup145 C) (26). Когда повторы CUG были экспрессированы в этом мутанте, сильный ядерный сигнал был обнаружен с помощью зонда CAG, но не с помощью зонда CTG (рис.А, нижняя часть). Напротив, ядерное накопление наблюдали только с зондом CTG, когда были экспрессированы повторы CAG (рис. A, верхняя часть). Это указывает на то, что зонды CNG могут проникать в ядро, специфичны для цепи и не гибридизуются с молекулами ДНК, содержащими тринуклеотидные повторы.
Способность зонда проникать в ядро оценивали на штамме, который, как известно, нарушает ядерный экспорт мРНК (Δ nup145 C) (26). Когда повторы CUG были экспрессированы в этом мутанте, сильный ядерный сигнал был обнаружен с помощью зонда CAG, но не с помощью зонда CTG (рис.А, нижняя часть). Напротив, ядерное накопление наблюдали только с зондом CTG, когда были экспрессированы повторы CAG (рис. A, верхняя часть). Это указывает на то, что зонды CNG могут проникать в ядро, специфичны для цепи и не гибридизуются с молекулами ДНК, содержащими тринуклеотидные повторы.
( A ) FISH-мечение в мутанте nup145C Δ, экспрессирующем либо URA3, -CAG 98 (вверху), либо URA3 -CUG 98 (внизу). мРНК детектировали с помощью олигонуклеотидных зондов, меченных CAG или CTG, и ядра окрашивали DAPI.( B ) Мечение FISH в штаммах, экспрессирующих URA3 (нет), URA3 -CAG 98 или URA3 -CUG 98 .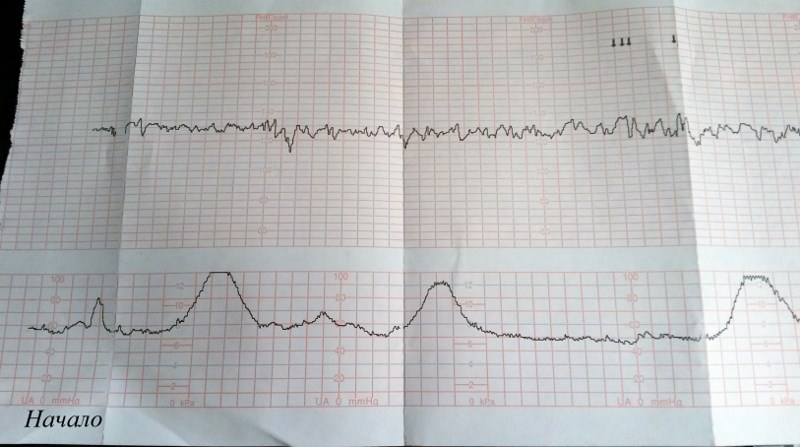 На вставках в правом нижнем углу панелей показаны части фигуры с большим увеличением. ( C ) Маркировка с помощью зонда олиго-dT.
На вставках в правом нижнем углу панелей показаны части фигуры с большим увеличением. ( C ) Маркировка с помощью зонда олиго-dT.
Мы гибридизировали штамм FYBL1-22B, трансформированный pE186, pTRI627 или pTRI628, с мечеными олигонуклеотидами CAG или CTG. В контрольном штамме (FYBL1-22B / pE186) при использовании обоих зондов было видно слабое окрашивание целых клеток, что соответствует фоновому уровню (рис.B, нет). В штамме FYBL1-22B / pTRI627, как и ожидалось, сигнал был виден только при зондировании олигонуклеотидом CTG, тогда как в FYBL1-22B / pTRI628 сигнал был обнаружен только с зондом CAG (рис. B, CAG 98). и CUG 98 ). Во всех меченых клетках мРНК, содержащие тринуклеотидный повтор, не были равномерно распределены по клетке, а образовывали агрегаты. Это неравномерное распределение не влияет на клеточные мРНК, которые не содержат таких повторов, поскольку те же штаммы, меченные олиго-dT, гибридизующимся с поли (A) -хвостом мРНК, не имеют этих кластеров (рис. C). Мутантный штамм Δ nup145 C показан в качестве положительного контроля для визуализации накопления мРНК в ядре и для оценки цитоплазматического фона. Мы также пытались проследить мРНК URA3 с хромосомными интегрированными конструкциями (GFR108 и GFR109), но сигналы были слишком слабыми, чтобы их можно было обнаружить. Чтобы обнаружить ядерное накопление транскриптов, ядерные сигналы в штаммах FYBL1-22B / pTRI627 и FYBL1-22B / pTRI628 были количественно определены и сравнены с сигналами, полученными в штамме, меченном олиго-dT зондом (см. Материалы и методы).Интенсивность ядерного окрашивания не увеличивается значительно в таких штаммах по сравнению с контрольными клетками, что позволяет предположить, что содержащие тринуклеотидные повторы мРНК специфически не накапливаются в ядерном компартменте. Мы пришли к выводу, что хотя транскрипты CAG и CUG образуют видимые внутриклеточные кластеры, явных дефектов ядерного экспорта нет.
C). Мутантный штамм Δ nup145 C показан в качестве положительного контроля для визуализации накопления мРНК в ядре и для оценки цитоплазматического фона. Мы также пытались проследить мРНК URA3 с хромосомными интегрированными конструкциями (GFR108 и GFR109), но сигналы были слишком слабыми, чтобы их можно было обнаружить. Чтобы обнаружить ядерное накопление транскриптов, ядерные сигналы в штаммах FYBL1-22B / pTRI627 и FYBL1-22B / pTRI628 были количественно определены и сравнены с сигналами, полученными в штамме, меченном олиго-dT зондом (см. Материалы и методы).Интенсивность ядерного окрашивания не увеличивается значительно в таких штаммах по сравнению с контрольными клетками, что позволяет предположить, что содержащие тринуклеотидные повторы мРНК специфически не накапливаются в ядерном компартменте. Мы пришли к выводу, что хотя транскрипты CAG и CUG образуют видимые внутриклеточные кластеры, явных дефектов ядерного экспорта нет.
ОБСУЖДЕНИЕ
В настоящей работе мы представляем доказательства того, что экспансии тринуклеотидных повторов происходят in vivo , в дрожжах, на уровне РНК, когда тринуклеотидные повторы CAG или CTG клонируются в 3′-UTR транскрибируемого гена Pol II.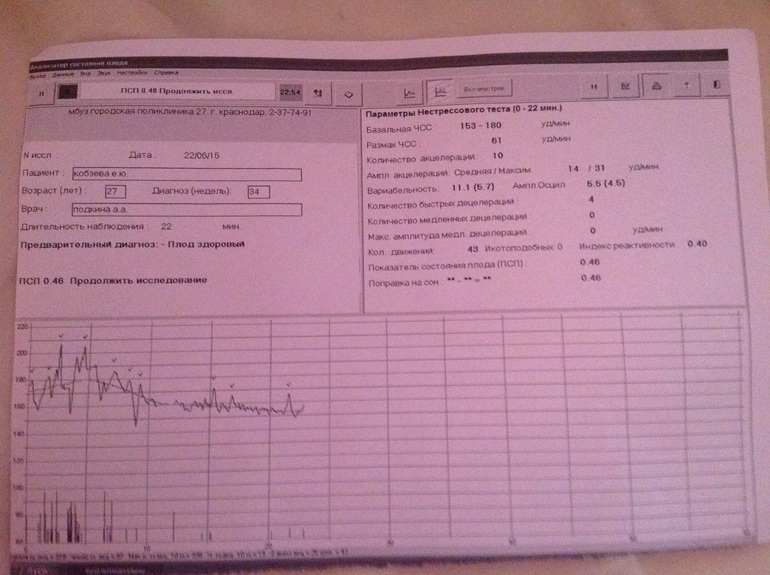 .Наблюдаются молекулы на 3 т.п.н. длиннее ожидаемого и заканчивающиеся на нормальном сайте терминатора транскрипции. Количество обнаруживаемой L-РНК выше, когда транскрибируются повторы CAG, что свидетельствует о зависимости от последовательности. Мы предполагаем, что эти большие молекулы РНК образуются за счет проскальзывания транскрипции, явления, которое, как сообщается, обычно стимулируется короткими мононуклеотидными или динуклеотидными повторами, как in vitro, (14–16,36), так и in vivo в E.coli. (17,18). Пациенты с болезнью Альцгеймера и Дауна обнаруживают проскальзывание транскрипции мРНК предшественника β-амилоида или мРНК убиквитина-B, что приводит к делециям моно- или динуклеотидного повтора внутри или рядом с повторяющимся участком (19, 20).Обычно предполагается, что во время транскрипции временная пауза в комплексе РНК-полимеразы способствует обратному проскальзыванию полимеразного комплекса, связанного с удлинением мРНК вдоль матрицы ДНК, и приводит к ресинтезу той же последовательности РНК (37).
.Наблюдаются молекулы на 3 т.п.н. длиннее ожидаемого и заканчивающиеся на нормальном сайте терминатора транскрипции. Количество обнаруживаемой L-РНК выше, когда транскрибируются повторы CAG, что свидетельствует о зависимости от последовательности. Мы предполагаем, что эти большие молекулы РНК образуются за счет проскальзывания транскрипции, явления, которое, как сообщается, обычно стимулируется короткими мононуклеотидными или динуклеотидными повторами, как in vitro, (14–16,36), так и in vivo в E.coli. (17,18). Пациенты с болезнью Альцгеймера и Дауна обнаруживают проскальзывание транскрипции мРНК предшественника β-амилоида или мРНК убиквитина-B, что приводит к делециям моно- или динуклеотидного повтора внутри или рядом с повторяющимся участком (19, 20).Обычно предполагается, что во время транскрипции временная пауза в комплексе РНК-полимеразы способствует обратному проскальзыванию полимеразного комплекса, связанного с удлинением мРНК вдоль матрицы ДНК, и приводит к ресинтезу той же последовательности РНК (37).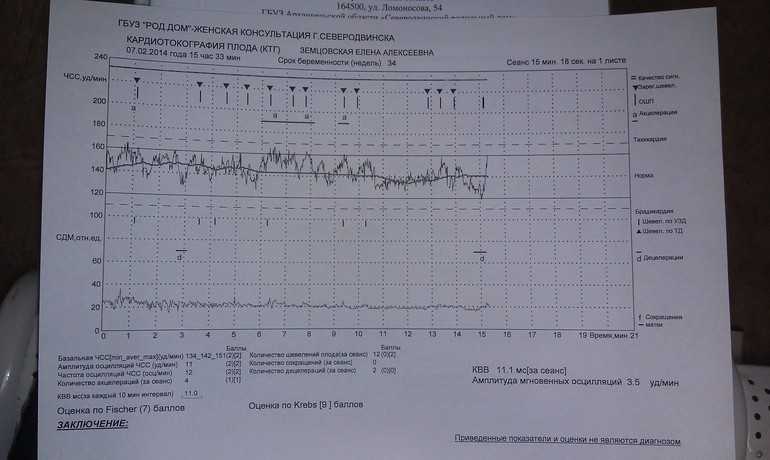 Тринуклеотидные повторы CAG / CTG образуют вторичные структуры in vitro (31–34), и было показано, что повторы CTG вызывают временные паузы в элонгации РНК после первого, второго и шестого-девятого триплетных повторов, in vitro ( 12). In vivo , CTG повторяет вилки остановки репликации в E.coli (38) и нарушает синтез ДНК, ассоциированной с рекомбинацией, в дрожжах (23). Мы наблюдали значительное снижение уровня L-РНК с конструкцией URA3 -TTG 87 (рис. E и A), подтверждая мнение о том, что стабильные вторичные структуры in vivo важны для проскальзывания транскрипции. Мы также обнаружили значительно более высокий уровень L-РНК с конструкцией URA3 -CTG 255 (рис.A), что снова согласуется с ролью вторичных структур в образовании L-РНК. Таким образом, мы предполагаем, что остановка комплекса РНК Pol II вторичными структурами, образованными на нити матричной ДНК, будет способствовать проскальзыванию и приведению к более длинным транскриптам, содержащим повторения повторяющейся последовательности.
Тринуклеотидные повторы CAG / CTG образуют вторичные структуры in vitro (31–34), и было показано, что повторы CTG вызывают временные паузы в элонгации РНК после первого, второго и шестого-девятого триплетных повторов, in vitro ( 12). In vivo , CTG повторяет вилки остановки репликации в E.coli (38) и нарушает синтез ДНК, ассоциированной с рекомбинацией, в дрожжах (23). Мы наблюдали значительное снижение уровня L-РНК с конструкцией URA3 -TTG 87 (рис. E и A), подтверждая мнение о том, что стабильные вторичные структуры in vivo важны для проскальзывания транскрипции. Мы также обнаружили значительно более высокий уровень L-РНК с конструкцией URA3 -CTG 255 (рис.A), что снова согласуется с ролью вторичных структур в образовании L-РНК. Таким образом, мы предполагаем, что остановка комплекса РНК Pol II вторичными структурами, образованными на нити матричной ДНК, будет способствовать проскальзыванию и приведению к более длинным транскриптам, содержащим повторения повторяющейся последовательности.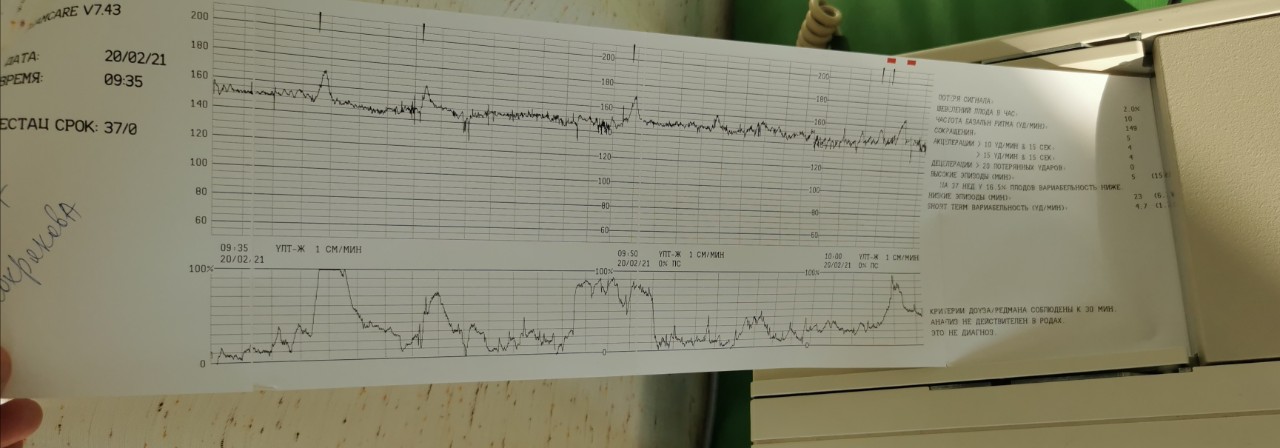 Вероятно, потребуется несколько раундов остановки / проскальзывания / синтеза для достижения размера обнаруженных L-РНК (Рис.). Если транскрипция не возобновляется после паузы, удлиняющаяся мРНК может деградировать, что приведет к снижению количества зрелой мРНК URA3 .Действительно, количество мРНК URA3 , обнаруженное в штамме GFR108 ( URA3 -CAG 98 ), составляло 40% от количества, обнаруженного в штамме GFR109 ( URA3 -CUG 98 ) (рис.). Однако мы не можем исключить возможность того, что относительная стабильность CAG- и CTG-содержащих транскриптов различна.
Вероятно, потребуется несколько раундов остановки / проскальзывания / синтеза для достижения размера обнаруженных L-РНК (Рис.). Если транскрипция не возобновляется после паузы, удлиняющаяся мРНК может деградировать, что приведет к снижению количества зрелой мРНК URA3 .Действительно, количество мРНК URA3 , обнаруженное в штамме GFR108 ( URA3 -CAG 98 ), составляло 40% от количества, обнаруженного в штамме GFR109 ( URA3 -CUG 98 ) (рис.). Однако мы не можем исключить возможность того, что относительная стабильность CAG- и CTG-содержащих транскриптов различна.
Интересно, что мы обнаружили больше L-РНК в штамме, который использует цепь CTG в качестве матрицы во время транскрипции, чем в штамме, использующем цепь CAG в качестве матрицы (рис.). Возможно, что вторичные структуры, образованные CTG-повторами in vivo , более стабильны, чем структуры, образованные CAG-повторами, что увеличивает вероятность приостановки полимеразы и, как следствие, проскальзывания транскрипции. Однако разница между повторами CAG и CTG уменьшалась, когда они переносились плазмидой с большим числом копий (рис. A), что позволяет предположить, что увеличение числа копий матричной ДНК изменяет величину проскальзывания или, альтернативно, что факторы хроматинизма являются по-разному собирается на плазмиде и на хромосоме, что может приводить к различным свойствам аппарата транскрипции.
Однако разница между повторами CAG и CTG уменьшалась, когда они переносились плазмидой с большим числом копий (рис. A), что позволяет предположить, что увеличение числа копий матричной ДНК изменяет величину проскальзывания или, альтернативно, что факторы хроматинизма являются по-разному собирается на плазмиде и на хромосоме, что может приводить к различным свойствам аппарата транскрипции.
Мы исследовали, подвержены ли естественные тринуклеотидные повторы в дрожжах проскальзыванию транскрипции. Не существует природной совершенной тринуклеотидной повторяющейся последовательности длины, сравнимой с CAG 98 у дрожжей (39). YPL009c содержит участок из 33 несовершенных тринуклеотидных повторов GAA (один из самых длинных тринуклеотидных повторов в геноме дрожжей), кодирующих глутаминовую кислоту или лизин. МРНК нормального размера, но не L-РНК, была обнаружена на нозерн-блотах для этого гена (данные не показаны), что позволяет предположить, что вырожденные повторы имеют тенденцию уменьшать проскальзывание, или что существует порог длины, ниже которого проскальзывание не может произойти, или что повторы GAA менее склонны к проскальзыванию.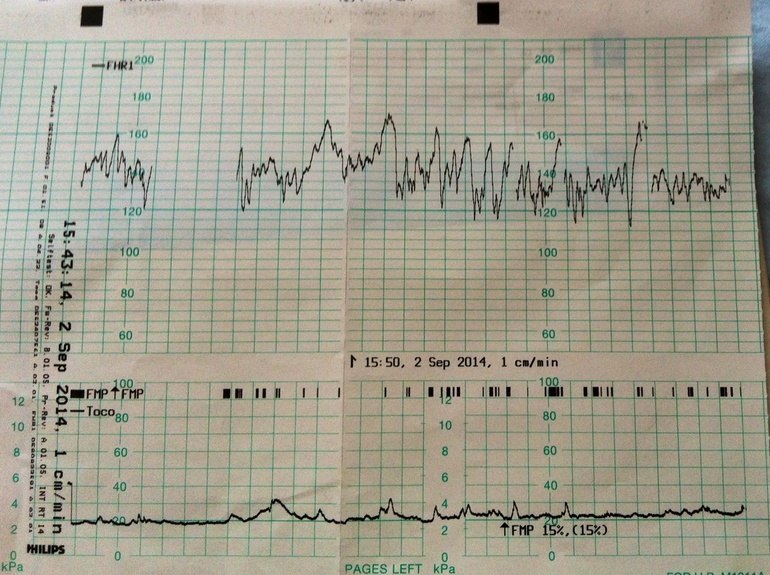 Эти три гипотезы не исключают друг друга.
Эти три гипотезы не исключают друг друга.
Проскальзывание транскрипции в тринуклеотидных повторах может также происходить в системах млекопитающих. Анализ CTG-содержащих транскриптов в нормальных и миотонических миобластах показал большие вариации размеров мРНК мутанта DMPK (8). Это было приписано гетерогенности размера повторов на уровне ДНК среди культивируемых фибробластов. Однако возможно, что некоторые из наблюдаемых различных молекул мРНК возникли в результате проскальзывания транскрипции.
В предыдущих исследованиях, в которых авторы изучали клеточное распределение транскриптов, содержащих длинные тринуклеотидные повторы, было показано, что хотя длинные транскрипты были правильно сплайсированы и полиаденилированы, они не были правильно экспортированы в цитоплазму. Сообщалось, что мРНК, содержащие CUG, накапливаются в ядрах миобластов, культивируемых при DM (7,8,11), и в ядрах мышечных клеток трансгенных мышей, экспрессирующих нетранслируемые повторы CUG (9).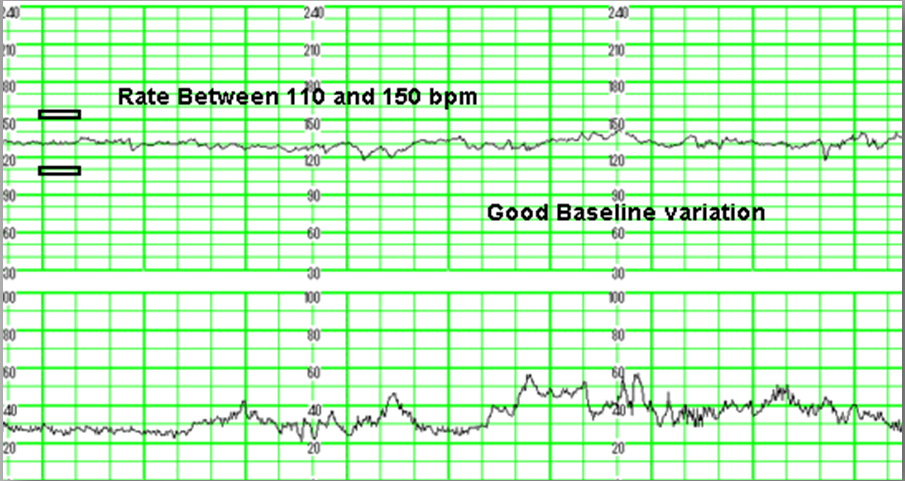 Совсем недавно было показано, что миотоническая дистрофия 2, вызванная экспансией тетрануклеотидного повтора CCTG в первом интроне гена ZNF9 , также обнаруживает ядерную задержку транскриптов (10).В дрожжах мы обнаружили, что L-РНК очищаются совместно с фракцией поли (A) + , что позволяет предположить, что они полиаденилированы, но CAG- и CUG-содержащие мРНК не обнаруживают каких-либо явных дефектов ядерного экспорта, хотя они имеют тенденцию к образуют кластеры в дрожжевых клетках, по-видимому, не специфичные для какого-либо субклеточного компартмента. Как CAG-, так и CUG-содержащие транскрипты агрегируются в дрожжах, тогда как у людей до сих пор были задокументированы только кластеры CUG-содержащих транскриптов. Какова природа этих кластеров? Было высказано предположение, что взаимодействия между расширенными повторами CUG и белками, связывающими двухцепочечную РНК, такими как гомологи белка Drosophila muscleblind или белка PKR (последний из которых имеет дрожжевые гомологи), могут объяснить ядерное удержание транскриптов мутантного DMPK (11,35).
Совсем недавно было показано, что миотоническая дистрофия 2, вызванная экспансией тетрануклеотидного повтора CCTG в первом интроне гена ZNF9 , также обнаруживает ядерную задержку транскриптов (10).В дрожжах мы обнаружили, что L-РНК очищаются совместно с фракцией поли (A) + , что позволяет предположить, что они полиаденилированы, но CAG- и CUG-содержащие мРНК не обнаруживают каких-либо явных дефектов ядерного экспорта, хотя они имеют тенденцию к образуют кластеры в дрожжевых клетках, по-видимому, не специфичные для какого-либо субклеточного компартмента. Как CAG-, так и CUG-содержащие транскрипты агрегируются в дрожжах, тогда как у людей до сих пор были задокументированы только кластеры CUG-содержащих транскриптов. Какова природа этих кластеров? Было высказано предположение, что взаимодействия между расширенными повторами CUG и белками, связывающими двухцепочечную РНК, такими как гомологи белка Drosophila muscleblind или белка PKR (последний из которых имеет дрожжевые гомологи), могут объяснить ядерное удержание транскриптов мутантного DMPK (11,35).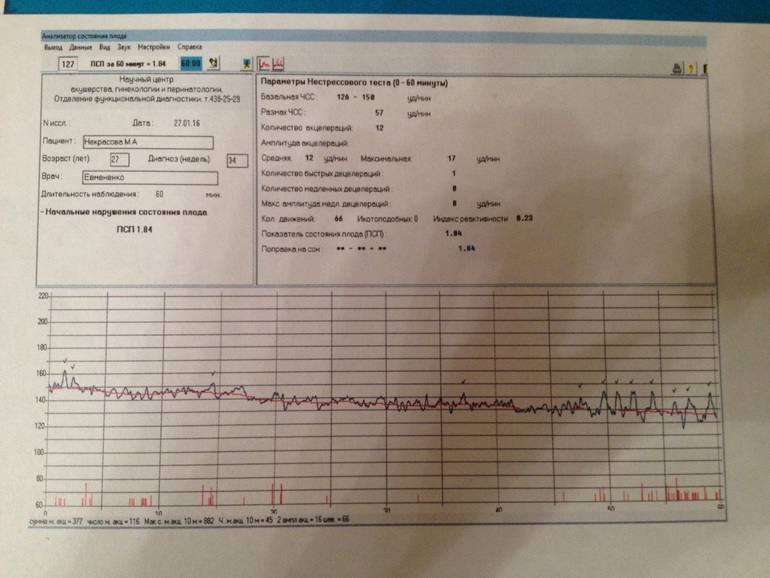 Альтернативно, они могут возникать, когда транскрипты, содержащие тринуклеотидные повторы, экспрессируются из-за межмолекулярных взаимодействий между такими РНК, причем обе гипотезы не исключают друг друга.
Альтернативно, они могут возникать, когда транскрипты, содержащие тринуклеотидные повторы, экспрессируются из-за межмолекулярных взаимодействий между такими РНК, причем обе гипотезы не исключают друг друга.
ДОПОЛНИТЕЛЬНЫЕ МАТЕРИАЛЫ
Дополнительные материалы доступны в NAR Online.
Дополнительные материалы доступны в NAR Online.
БЛАГОДАРНОСТИ
Мы благодарим J. Boyer за щедрый дар плазмиды pCMha191, M. Richetti и G. Fischer за критическое прочтение рукописи и наших коллег за плодотворные обсуждения.Мы также благодарим B. Arcangioli и A. Jacquier за соответствующие технические советы и вдумчивые комментарии к настоящей работе. B.D. является членом Institut Universitaire de France.
ССЫЛКИ
1. Каммингс К.Дж. и Зогби, Х.Й. (2000) Четырнадцать и их количество: раскрытие болезней с тринуклеотидными повторами. Гм. Мол. Genet., 9, 909–916. [PubMed] [Google Scholar] 2. Альба М.М., Сантибанес-Кореф М.Ф. и Хэнкок, Дж. (1999) Повторы аминокислот в дрожжах чрезмерно представлены в определенных классах белков и демонстрируют свидетельства мутационного процесса, подобного проскальзыванию. J. Mol. Evol., 49, 789–797. [PubMed] [Google Scholar] 3. Брук Дж. Д., МакКеррач, М. Э., Харли, Х. Дж., Баклер, А. Дж., Чёрч, Д., Абуратани, Г., Хантер, К., Стэнтон, В. П., Тирион, Д. П., Хадсон, Т. и др. . (1992) Молекулярная основа миотонической дистрофии: расширение тринуклеотидного (CTG) повтора на 3′-конце транскрипта, кодирующего член семейства протеинкиназ. Cell, 68, 799–808. [PubMed] [Google Scholar] 4. Харли Х. Г., Брук, Д. Д., Рандл, С. А., Кроу, С., Рирдон, В., Баклер, А. Дж., Харпер, П. С., Хаусман, Д.Э. и Шоу Д. (1992) Расширение нестабильной области ДНК и фенотипические вариации при миотонической дистрофии. Природа, 355, 545–546. [PubMed] [Google Scholar] 5. Куб М.Д., Мозли М.Л., Шут Л.Дж., Бенцов К.А., Берд Т.Д., Дэй, Дж. У. и Ранум, L.P.W. (1999) Нетранслируемая экспансия CTG вызывает новую форму спиноцеребеллярной атаксии (SCA8). Nature Genet., 21, 379–384. [PubMed] [Google Scholar] 6. Krahe R., Ashizawa, T., Abbruzzese, C., Roeder, E., Carango, P., Giacanelli, M.
J. Mol. Evol., 49, 789–797. [PubMed] [Google Scholar] 3. Брук Дж. Д., МакКеррач, М. Э., Харли, Х. Дж., Баклер, А. Дж., Чёрч, Д., Абуратани, Г., Хантер, К., Стэнтон, В. П., Тирион, Д. П., Хадсон, Т. и др. . (1992) Молекулярная основа миотонической дистрофии: расширение тринуклеотидного (CTG) повтора на 3′-конце транскрипта, кодирующего член семейства протеинкиназ. Cell, 68, 799–808. [PubMed] [Google Scholar] 4. Харли Х. Г., Брук, Д. Д., Рандл, С. А., Кроу, С., Рирдон, В., Баклер, А. Дж., Харпер, П. С., Хаусман, Д.Э. и Шоу Д. (1992) Расширение нестабильной области ДНК и фенотипические вариации при миотонической дистрофии. Природа, 355, 545–546. [PubMed] [Google Scholar] 5. Куб М.Д., Мозли М.Л., Шут Л.Дж., Бенцов К.А., Берд Т.Д., Дэй, Дж. У. и Ранум, L.P.W. (1999) Нетранслируемая экспансия CTG вызывает новую форму спиноцеребеллярной атаксии (SCA8). Nature Genet., 21, 379–384. [PubMed] [Google Scholar] 6. Krahe R., Ashizawa, T., Abbruzzese, C., Roeder, E., Carango, P., Giacanelli, M. , Funanage, V.L. и Сицилиано М.Дж. (1995) Эффект экспансии тринуклеотидного повтора миотонической дистрофии на транскрипцию и процессинг DMPK .Геномика, 28, 1–14. [PubMed] [Google Scholar] 7. Танежа К.Л., МакКеррач М., Шаллинг М., Хаусман Д. и Сингер Р. (1995) Очаги транскриптов тринуклеотидных повторов в ядрах клеток и тканей миотонической дистрофии. J. Cell Biol., 128, 995–1002. [Бесплатная статья PMC] [PubMed] [Google Scholar] 8. Дэвис Б.М., МакКеррач М.Э., Танеха К.Л., Сингер Р.Х. и Housman, D.E. (1997) Экспансия тринуклеотидного повтора CUG в 3′-нетранслируемой области транскриптов протеинкиназы миотонической дистрофии приводит к сохранению транскриптов в ядре.Proc. Natl Acad. Sci. США, 94, 7388–7393. [Бесплатная статья PMC] [PubMed] [Google Scholar] 9. Манкоди А., Логиджиан Э., Каллахан Л., Макклейн Ч., Уайт Р., Хендерсон Д., Крым М. и Торнтон, К.А. (2000) Миотоническая дистрофия у трансгенных мышей, экспрессирующих увеличенный повтор CUG. Science, 289, 1769–1772. [PubMed] [Google Scholar] 10.
, Funanage, V.L. и Сицилиано М.Дж. (1995) Эффект экспансии тринуклеотидного повтора миотонической дистрофии на транскрипцию и процессинг DMPK .Геномика, 28, 1–14. [PubMed] [Google Scholar] 7. Танежа К.Л., МакКеррач М., Шаллинг М., Хаусман Д. и Сингер Р. (1995) Очаги транскриптов тринуклеотидных повторов в ядрах клеток и тканей миотонической дистрофии. J. Cell Biol., 128, 995–1002. [Бесплатная статья PMC] [PubMed] [Google Scholar] 8. Дэвис Б.М., МакКеррач М.Э., Танеха К.Л., Сингер Р.Х. и Housman, D.E. (1997) Экспансия тринуклеотидного повтора CUG в 3′-нетранслируемой области транскриптов протеинкиназы миотонической дистрофии приводит к сохранению транскриптов в ядре.Proc. Natl Acad. Sci. США, 94, 7388–7393. [Бесплатная статья PMC] [PubMed] [Google Scholar] 9. Манкоди А., Логиджиан Э., Каллахан Л., Макклейн Ч., Уайт Р., Хендерсон Д., Крым М. и Торнтон, К.А. (2000) Миотоническая дистрофия у трансгенных мышей, экспрессирующих увеличенный повтор CUG. Science, 289, 1769–1772. [PubMed] [Google Scholar] 10.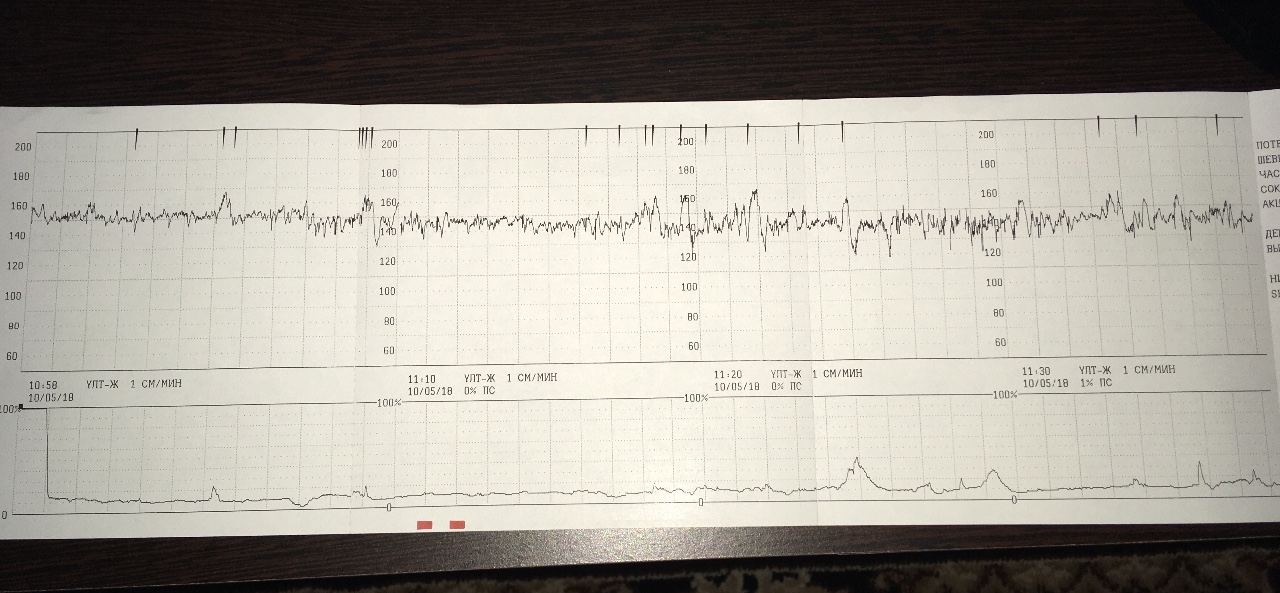 Ликури К.Л., Рикер К., Мозли М.Л., Якобсен Дж.Ф., Кресс В., Нейлор С.Л., Дэй, Дж. У. и Ранум, L.P.W. (2001) Миотоническая дистрофия 2 типа, вызванная экспансией CCTG в интроне 1 ZNF9 .Science, 293, 864–867. [PubMed] [Google Scholar] 11. Миллер Дж. У., Урбинати К. Р., Тенгумнуай П., Стенберг М. Г., Бирн Б. Дж., Торнтон К. А. и Swanson, M.S. (2000) Привлечение человеческих белков, связывающих мышцы, в экспансию (CUG) n, связанную с миотонической дистрофией. EMBO J., 19, 4439–4448. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Парсонс М.А., Синден Р.Р. и Избань М. (1998) Транскрипционные свойства РНК-полимеразы II в ДНК, содержащей триплетные повторы, из миотонической дистрофии человека и ломких Х-локусов.J. Biol. Chem., 273, 26998–27008. [PubMed] [Google Scholar] 13. Макдональд Дж. Ф. (1993) Эволюция и последствия мобильных элементов. Curr. Opin. Genet. Дев., 3, 855–864. [PubMed] [Google Scholar] 14. Северинов К., Гольдфарб А. (1994) Топология сайта связывания продукта в РНК-полимеразе, выявленная по проскальзыванию транскрипта на промоторе фага P L .
Ликури К.Л., Рикер К., Мозли М.Л., Якобсен Дж.Ф., Кресс В., Нейлор С.Л., Дэй, Дж. У. и Ранум, L.P.W. (2001) Миотоническая дистрофия 2 типа, вызванная экспансией CCTG в интроне 1 ZNF9 .Science, 293, 864–867. [PubMed] [Google Scholar] 11. Миллер Дж. У., Урбинати К. Р., Тенгумнуай П., Стенберг М. Г., Бирн Б. Дж., Торнтон К. А. и Swanson, M.S. (2000) Привлечение человеческих белков, связывающих мышцы, в экспансию (CUG) n, связанную с миотонической дистрофией. EMBO J., 19, 4439–4448. [Бесплатная статья PMC] [PubMed] [Google Scholar] 12. Парсонс М.А., Синден Р.Р. и Избань М. (1998) Транскрипционные свойства РНК-полимеразы II в ДНК, содержащей триплетные повторы, из миотонической дистрофии человека и ломких Х-локусов.J. Biol. Chem., 273, 26998–27008. [PubMed] [Google Scholar] 13. Макдональд Дж. Ф. (1993) Эволюция и последствия мобильных элементов. Curr. Opin. Genet. Дев., 3, 855–864. [PubMed] [Google Scholar] 14. Северинов К., Гольдфарб А. (1994) Топология сайта связывания продукта в РНК-полимеразе, выявленная по проскальзыванию транскрипта на промоторе фага P L . J. Biol. Chem., 269, 31701–31705. [PubMed] [Google Scholar] 15. Jeong S.W., Lang, W.H. и Reeder, R.H. (1996) Дрожжевой терминатор транскрипции для РНК-полимеразы I разработан для предотвращения проскальзывания полимеразы.J. Biol. Chem., 271, 16104–16110. [PubMed] [Google Scholar] 16. Дэн Л., Шуман С. (1997) Свойства удлинения РНК-полимеразы вируса осповакцины: пауза, проскальзывание, добавление 3′-конца и выбор сайта терминации. Биохимия, 36, 15892–15899. [PubMed] [Google Scholar] 17. Вагнер Л.А., Вайс Р.Б., Дрисколл Р., Данн Д.С. и Гестеланд Р.Ф. (1990) Транскрипционное проскальзывание происходит во время удлинения при прохождении аденина или тимина в Escherichia coli . Nucleic Acids Res., 18, 3529–3535. [Бесплатная статья PMC] [PubMed] [Google Scholar] 18.Сюн X.F. и Резникофф, В. (1993) Проскальзывание транскрипции во время процесса инициации транскрипции на мутантном промоторе lac in vivo . J. Mol. Биол., 231, 569–580. [PubMed] [Google Scholar] 19. van Leeuwen F.
J. Biol. Chem., 269, 31701–31705. [PubMed] [Google Scholar] 15. Jeong S.W., Lang, W.H. и Reeder, R.H. (1996) Дрожжевой терминатор транскрипции для РНК-полимеразы I разработан для предотвращения проскальзывания полимеразы.J. Biol. Chem., 271, 16104–16110. [PubMed] [Google Scholar] 16. Дэн Л., Шуман С. (1997) Свойства удлинения РНК-полимеразы вируса осповакцины: пауза, проскальзывание, добавление 3′-конца и выбор сайта терминации. Биохимия, 36, 15892–15899. [PubMed] [Google Scholar] 17. Вагнер Л.А., Вайс Р.Б., Дрисколл Р., Данн Д.С. и Гестеланд Р.Ф. (1990) Транскрипционное проскальзывание происходит во время удлинения при прохождении аденина или тимина в Escherichia coli . Nucleic Acids Res., 18, 3529–3535. [Бесплатная статья PMC] [PubMed] [Google Scholar] 18.Сюн X.F. и Резникофф, В. (1993) Проскальзывание транскрипции во время процесса инициации транскрипции на мутантном промоторе lac in vivo . J. Mol. Биол., 231, 569–580. [PubMed] [Google Scholar] 19. van Leeuwen F. W., де Клейн, D.P.V., van den Hurk, H.H., Neubauer, A., Sonnemans, M.A.F., Sluijs, J.A., Köycü, S., Ramdjielal, R.D.J., Salehi, A., Martens, G.J.M. и др. . (1998) Мутанты смещения рамки считывания белка-предшественника β-амилоида и убиквитина-B у пациентов с болезнью Альцгеймера и Дауна.Наука, 279, 242–247. [PubMed] [Google Scholar] 20. van den Hurk W.H., Willems, H.J.J., Bloemen, M. и Martens, G.J.M. (2001) Новые мутации сдвига рамки считывания около коротких простых повторов. J. Biol. Chem., 276, 11496–11498. [PubMed] [Google Scholar] 21. Гари Э., Пьедрафита Л., Алдеа М. и Эрреро, Э. (1997) Набор векторов с регулируемой тетрациклином промоторной системой для модуляции экспрессии генов в Saccharomyces cerevisiae . Дрожжи, 13, 837–848. [PubMed] [Google Scholar] 22. Канг С., Яворски А., Ohshima, K. и Уэллс, Р. (1995) Экспансия и делеция повторов CTG из генов болезней человека определяются направлением репликации в E. coli . Nature Genet., 10, 213–217. [PubMed] [Google Scholar] 23.
W., де Клейн, D.P.V., van den Hurk, H.H., Neubauer, A., Sonnemans, M.A.F., Sluijs, J.A., Köycü, S., Ramdjielal, R.D.J., Salehi, A., Martens, G.J.M. и др. . (1998) Мутанты смещения рамки считывания белка-предшественника β-амилоида и убиквитина-B у пациентов с болезнью Альцгеймера и Дауна.Наука, 279, 242–247. [PubMed] [Google Scholar] 20. van den Hurk W.H., Willems, H.J.J., Bloemen, M. и Martens, G.J.M. (2001) Новые мутации сдвига рамки считывания около коротких простых повторов. J. Biol. Chem., 276, 11496–11498. [PubMed] [Google Scholar] 21. Гари Э., Пьедрафита Л., Алдеа М. и Эрреро, Э. (1997) Набор векторов с регулируемой тетрациклином промоторной системой для модуляции экспрессии генов в Saccharomyces cerevisiae . Дрожжи, 13, 837–848. [PubMed] [Google Scholar] 22. Канг С., Яворски А., Ohshima, K. и Уэллс, Р. (1995) Экспансия и делеция повторов CTG из генов болезней человека определяются направлением репликации в E. coli . Nature Genet., 10, 213–217. [PubMed] [Google Scholar] 23. Ричард Г.-Ф., Гёльнер Г.М., МакМюррей, К.Т. и Хабер, Дж. (2000) Индуцированные рекомбинацией экспансии тринуклеотидных повторов CAG в дрожжах включают комплекс MRE11, / RAD50, /, XRS2, . EMBO J., 19, 2381–2390. [Бесплатная статья PMC] [PubMed] [Google Scholar] 24.Фэйрхед К., Льоренте Б., Денис Ф., Солер М. и Дуйон, Б. (1996) Новые векторы для комбинаторных делеций в хромосомах дрожжей и для клонирования с восстановлением разрывов с использованием рекомбинации «маркеры расщепления». Дрожжи, 12, 1439–1457. [PubMed] [Google Scholar] 25. Чен Д.-К., Ян Б.-К. и Куо, Т.-Т. (1992) Одностадийная трансформация дрожжей в стационарной фазе. Curr. Genet., 21, 83–84. [PubMed] [Google Scholar] 26. Тейшейра М.Т., Синиоссоглу С., Подтелейников С., Бенишоу Ж.-К., Манн М., Дуйон Б., Хёрт Э. и Фабр, Э. (1997) Два функционально различных домена, генерируемых in vivo расщеплением Nup145p: новый путь биогенеза нуклеопоринов.EMBO J., 16, 5086–5097. [Бесплатная статья PMC] [PubMed] [Google Scholar] 27.
Ричард Г.-Ф., Гёльнер Г.М., МакМюррей, К.Т. и Хабер, Дж. (2000) Индуцированные рекомбинацией экспансии тринуклеотидных повторов CAG в дрожжах включают комплекс MRE11, / RAD50, /, XRS2, . EMBO J., 19, 2381–2390. [Бесплатная статья PMC] [PubMed] [Google Scholar] 24.Фэйрхед К., Льоренте Б., Денис Ф., Солер М. и Дуйон, Б. (1996) Новые векторы для комбинаторных делеций в хромосомах дрожжей и для клонирования с восстановлением разрывов с использованием рекомбинации «маркеры расщепления». Дрожжи, 12, 1439–1457. [PubMed] [Google Scholar] 25. Чен Д.-К., Ян Б.-К. и Куо, Т.-Т. (1992) Одностадийная трансформация дрожжей в стационарной фазе. Curr. Genet., 21, 83–84. [PubMed] [Google Scholar] 26. Тейшейра М.Т., Синиоссоглу С., Подтелейников С., Бенишоу Ж.-К., Манн М., Дуйон Б., Хёрт Э. и Фабр, Э. (1997) Два функционально различных домена, генерируемых in vivo расщеплением Nup145p: новый путь биогенеза нуклеопоринов.EMBO J., 16, 5086–5097. [Бесплатная статья PMC] [PubMed] [Google Scholar] 27. Гитц Р.Д., Шистл Р.Х., Виллемс А.Р. и Вудс Р.А. (1995) Исследования трансформации интактных дрожжевых клеток с помощью процедуры LiAc / SS-ДНК / PEG. Дрожжи, 11, 355–360. [PubMed] [Google Scholar] 28. Ричард Г.-Ф., Фэрхед, К. и Дуйон, Б. (1997) Полная транскрипционная карта хромосомы XI дрожжей в различных условиях жизни. J. Mol. Биол., 268, 303–321. [PubMed] [Google Scholar] 29. Дженсен Т.Х., Патрисио, К., Маккарти, Т. и Росбаш М.(2001) Блокирование экспорта мРНК в ядро в S. cerevisiae приводит к гипераденилированию транскриптов, которые накапливаются в месте транскрипции. Мол. Cell, 7, 887–898. [PubMed] [Google Scholar] 30. Фэйрхед К. и Дуйон Б. (1994) Карта транскриптов двух участков хромосомы XI Saccharomyces cerevisiae для интерпретации результатов систематического секвенирования. Дрожжи, 10, 1403–1413. [PubMed] [Google Scholar] 31. Гейси А.М., Гёльнер Г., Юраник Н., Макура С. и МакМюррей, C.T. (1995) Тринуклеотидные повторы, которые расширяются при заболеваниях человека, образуют шпильки in vitro .
Гитц Р.Д., Шистл Р.Х., Виллемс А.Р. и Вудс Р.А. (1995) Исследования трансформации интактных дрожжевых клеток с помощью процедуры LiAc / SS-ДНК / PEG. Дрожжи, 11, 355–360. [PubMed] [Google Scholar] 28. Ричард Г.-Ф., Фэрхед, К. и Дуйон, Б. (1997) Полная транскрипционная карта хромосомы XI дрожжей в различных условиях жизни. J. Mol. Биол., 268, 303–321. [PubMed] [Google Scholar] 29. Дженсен Т.Х., Патрисио, К., Маккарти, Т. и Росбаш М.(2001) Блокирование экспорта мРНК в ядро в S. cerevisiae приводит к гипераденилированию транскриптов, которые накапливаются в месте транскрипции. Мол. Cell, 7, 887–898. [PubMed] [Google Scholar] 30. Фэйрхед К. и Дуйон Б. (1994) Карта транскриптов двух участков хромосомы XI Saccharomyces cerevisiae для интерпретации результатов систематического секвенирования. Дрожжи, 10, 1403–1413. [PubMed] [Google Scholar] 31. Гейси А.М., Гёльнер Г., Юраник Н., Макура С. и МакМюррей, C.T. (1995) Тринуклеотидные повторы, которые расширяются при заболеваниях человека, образуют шпильки in vitro . Cell, 81, 533–540. [PubMed] [Google Scholar] 32. Ю.А., Митас М. (1995) Богатые пуринами последовательности тринуклеотидных повторов d (CAG) 15 и d (GAC) 15 образуют шпильки. Nucleic Acids Res., 23, 4055–4057. [Бесплатная статья PMC] [PubMed] [Google Scholar] 33. Митас М., Ю. А., Дилл, Дж., Камп, Т. Дж., Чемберс, Э. Дж. и Хаворт, И.С. (1995) Шпильковые свойства одноцепочечной ДНК, содержащей GC-богатый триплетный повтор: (CTG) 15. Nucleic Acids Res., 23, 1050–1059. [Бесплатная статья PMC] [PubMed] [Google Scholar] 35. Тиан Б., Уайт, Р.Дж., Ся, Т., Велл, С., Тернер, Д.Х., Мэтьюз, М. и Торнтон, К.А. (2000) Расширенные РНК повторов CUG образуют шпильки, которые активируют двухцепочечную РНК-зависимую протеинкиназу PKR. РНК, 6, 79–87. [Бесплатная статья PMC] [PubMed] [Google Scholar] 36. Макдональд Л.Э., Чжоу Ю. и Макаллистер, W.T. (1993) Окончание и проскальзывание РНК-полимеразой бактериофага Т7. J. Mol. Биол., 232, 1030–1047. [PubMed] [Google Scholar] 37. Жак Ж.-П. и Колаковский Д. (1991) Псевдошаблонная транскрипция в прокариотических и эукариотических организмах.
Cell, 81, 533–540. [PubMed] [Google Scholar] 32. Ю.А., Митас М. (1995) Богатые пуринами последовательности тринуклеотидных повторов d (CAG) 15 и d (GAC) 15 образуют шпильки. Nucleic Acids Res., 23, 4055–4057. [Бесплатная статья PMC] [PubMed] [Google Scholar] 33. Митас М., Ю. А., Дилл, Дж., Камп, Т. Дж., Чемберс, Э. Дж. и Хаворт, И.С. (1995) Шпильковые свойства одноцепочечной ДНК, содержащей GC-богатый триплетный повтор: (CTG) 15. Nucleic Acids Res., 23, 1050–1059. [Бесплатная статья PMC] [PubMed] [Google Scholar] 35. Тиан Б., Уайт, Р.Дж., Ся, Т., Велл, С., Тернер, Д.Х., Мэтьюз, М. и Торнтон, К.А. (2000) Расширенные РНК повторов CUG образуют шпильки, которые активируют двухцепочечную РНК-зависимую протеинкиназу PKR. РНК, 6, 79–87. [Бесплатная статья PMC] [PubMed] [Google Scholar] 36. Макдональд Л.Э., Чжоу Ю. и Макаллистер, W.T. (1993) Окончание и проскальзывание РНК-полимеразой бактериофага Т7. J. Mol. Биол., 232, 1030–1047. [PubMed] [Google Scholar] 37. Жак Ж.-П. и Колаковский Д. (1991) Псевдошаблонная транскрипция в прокариотических и эукариотических организмах. Genes Dev., 5, 707–713. [PubMed] [Google Scholar] 38. Самадашвили Г., Раца Г. и Миркин С. (1997) Тринуклеотидные повторы влияют на репликацию ДНК in vivo . Nature Genet., 17, 298–304. [PubMed] [Google Scholar] 39. Ричард Г.-Ф. и Дуйон, Б. (1997) Тринуклеотидные повторы в дрожжах. Res. Microbiol., 148, 731–744. [PubMed] [Google Scholar] 40. Кумадзава Ю., Йокогава, Т., Цуруи, Х., Миура, К.-Я. и Ватанабэ, К. (1992) Влияние структуры тРНК высшего порядка на стабильность гибридов с олигодезоксирибонуклеотидами: разделение тРНК с помощью эффективной гибридизации в растворе Nucleic Acids Res., 20, 2223–2232. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Genes Dev., 5, 707–713. [PubMed] [Google Scholar] 38. Самадашвили Г., Раца Г. и Миркин С. (1997) Тринуклеотидные повторы влияют на репликацию ДНК in vivo . Nature Genet., 17, 298–304. [PubMed] [Google Scholar] 39. Ричард Г.-Ф. и Дуйон, Б. (1997) Тринуклеотидные повторы в дрожжах. Res. Microbiol., 148, 731–744. [PubMed] [Google Scholar] 40. Кумадзава Ю., Йокогава, Т., Цуруи, Х., Миура, К.-Я. и Ватанабэ, К. (1992) Влияние структуры тРНК высшего порядка на стабильность гибридов с олигодезоксирибонуклеотидами: разделение тРНК с помощью эффективной гибридизации в растворе Nucleic Acids Res., 20, 2223–2232. [Бесплатная статья PMC] [PubMed] [Google Scholar]Экспансия CTG-повтора Фукса, связанная с дистрофией роговицы, в гене TCF4 влияет на транскрипцию с его альтернативных промоторов
Zhuang, Y., Cheng, P. & Weintraub, H Развитие В-лимфоцитов регулируется комбинированной дозой трех основных генов спираль-петля-спираль, E2A, E2–2 и HEB. Мол. Клетка. Биол. 16 , 2898–2905 (1996).
CAS Статья Google Scholar
Guillemot, F. Пространственная и временная спецификация нервных судеб с помощью кодов факторов транскрипции. Разработка 134 , 3771–3780 (2007).
CAS Статья Google Scholar
Zweier, C. et al. Гаплонедостаточность TCF4 вызывает синдромальную умственную отсталость с перемежающейся гипервентиляцией (синдром Питта-Хопкинса). Am. J. Hum. Genet. 80 , 994–1001 (2007).
CAS Статья Google Scholar
Sepp, M., Kannike, K., Eesmaa, A., Urb, M. & Timmusk, T. Функциональное разнообразие изоформ человеческого фактора транскрипции TCF4 основного спираль-петля-спираль, генерируемых альтернативным 5′-экзоном использование и сращивание. PLoS ONE 6 , e22138 (2011).
ADS CAS Статья Google Scholar

Фагерберг, Л. et al. Анализ тканеспецифической экспрессии человека путем полногеномной интеграции транскриптомики и протеомики на основе антител. Мол. Cell Proteomics 13 , 397–406 (2014).
CAS Статья Google Scholar
Ранналс, М. Д. и Махер, Б. Дж. Молекулярные механизмы фактора транскрипции 4 при синдроме Питта Хопкинса. Curr. Genet. Med. Отчет 5 , 1–7 (2017).
Артикул Google Scholar
Ma, C., Gu, C., Huo, Y., Li, X. & Luo, X.-J. Интегрированный ландшафт причинных генов и путей при шизофрении. Пер. Психиатрия https://doi.org/10.1038/s41398-018-0114-x (2018).
Артикул PubMed PubMed Central Google Scholar
Sepp, M. et al. Фактор транскрипции TCF4, связанный с умственной отсталостью и шизофренией, регулируется нейрональной активностью и протеинкиназой А. J. Neurosci. 37 , 10516–10527 (2017).
J. Neurosci. 37 , 10516–10527 (2017).
CAS Статья Google Scholar
Ripke, S. et al. Биологические выводы из 108 генетических локусов, связанных с шизофренией. Nature 511 , 421–427 (2014).
ADS CAS Google Scholar
Stefansson, H. et al. Общие варианты, приводящие к риску шизофрении. Nature 460 , 744–747 (2009).
ADS CAS Статья Google Scholar
Doostparast Torshizi, A. et al. Деконволюция транскрипционных сетей идентифицирует TCF4 как главный регулятор при шизофрении. Sci. Adv. 5 , 4139 (2019).
ADS Статья Google Scholar
Харбанда, М. et al. Частичная делеция TCF4 в семье из трех поколений с несиндромальной интеллектуальной инвалидностью, без признаков синдрома Питта-Хопкинса.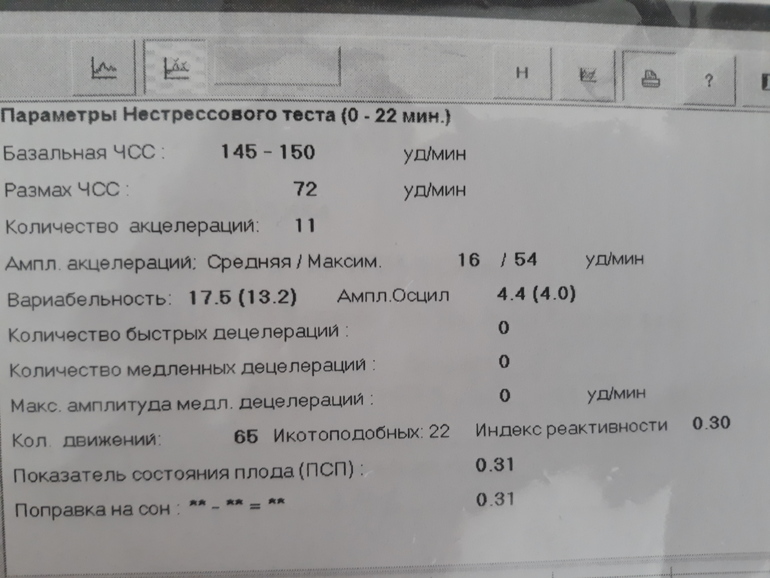 евро. J. Med. Genet. 59 , 310–314 (2016).
евро. J. Med. Genet. 59 , 310–314 (2016).
Артикул Google Scholar
Wray, N. R. et al. Полногеномный ассоциативный анализ выявляет 44 варианта риска и уточняет генетическую архитектуру большой депрессии. Нат. Genet. 50 , 668 (2018).
CAS Статья Google Scholar
Wieben, E. D. et al. Распространение распространенного тринуклеотидного повтора в гене фактора транскрипции 4 (TCF4, E2–2) предсказывает дистрофию роговицы Фукса. PLoS ONE 7 , e49083 (2012).
ADS CAS Статья Google Scholar
Дель-Фаверо, Дж. et al. Европейский комбинированный анализ повторов CTG18.1 и ERDA1 CAG / CTG при биполярном расстройстве. евро. J. Hum. Genet. 10 , 276–280 (2002).
CAS Статья Google Scholar

Fautsch, M. P. et al. TCF4-опосредованная эндотелиальная дистрофия роговицы Фукса: понимание распространенного заболевания, связанного с тринуклеотидными повторами. Прог. Retinal Eye Res. https: // doi.org / 10.1016 / j.preteyeres.2020.100883 (2020).
Артикул Google Scholar
Ong Tone, S. et al. Эндотелиальная дистрофия роговицы Фукса: порочный круг патогенеза Фукса. Прог. Retinal Eye Res. https://doi.org/10.1016/j.preteyeres.2020.100863 (2020).
Артикул Google Scholar
Vieta, E. et al. Биполярные расстройства. Нат. Преподобный Дис. Праймеры 4 , 1–16 (2018).
Артикул Google Scholar
Foja, S., Luther, M., Hoffmann, K., Rupprecht, A. & Gruenauer-Kloevekorn, C. Экспансия повтора CTG181 может снижать экспрессию гена TCF4 в эндотелиальных клетках роговицы немецких пациентов с синдромом Фукса. дистрофия. Graefes Arch. Clin. Exp. Офтальмол. 255 , 1621–1631 (2017).
дистрофия. Graefes Arch. Clin. Exp. Офтальмол. 255 , 1621–1631 (2017).
CAS Статья Google Scholar
Okumura, N. et al. Влияние экспансии тринуклеотидных повторов на экспрессию мРНК TCF4 при эндотелиальной дистрофии роговицы Фукса. Investig. Офтальмол. Vis. Sci. 60 , 779–786 (2019).
CAS Статья Google Scholar
Mootha, V. V. et al. Экспансия триплетного повтора TCF4 и фокусы ядерной РНК при эндотелиальной дистрофии роговицы Фукса. Investig. Офтальмол.Vis. Sci. 56 , 2003–2011 (2015).
CAS Статья Google Scholar
Ołdak, M. et al. Эндотелиальная дистрофия роговицы по Фуксу: сильная ассоциация с rs613872, не сопровождающаяся изменениями уровня мРНК TCF4 в эндотелиальной оболочке роговицы. Biomed. Res. Int. 2015 , 640234 (2015).
Res. Int. 2015 , 640234 (2015).
Артикул Google Scholar
Уорд, М. С. и Гилад, Ю. Геномика человека: взлом регуляторного кода. Nature 550 , 190–191 (2017).
ADS Статья Google Scholar
Ювен-Гершон, Т., Хсу, Дж.-Й., Тайзен, Дж. У. М., Кадонага, Дж. Т. Основной промотор РНК-полимеразы II — путь к транскрипции. Curr. Opin. Cell Biol. 20 , 253–259 (2008).
CAS Статья Google Scholar
Brandenberger, R. et al. Характеристика транскриптома проясняет сигнальные сети, которые контролируют рост и дифференцировку ES-клеток человека. Нат. Biotechnol. 22 , 707–716 (2004).
Артикул Google Scholar
Suzuki, Y. et al. Крупномасштабный сбор и характеристика промоторов генов человека и мыши. In Silico Biol. 4 , 429–444 (2004).
In Silico Biol. 4 , 429–444 (2004).
PubMed Google Scholar
Никитина А.С. и др. Набор данных по транскриптомному профилированию эндотелия роговицы пациентов с эндотелиальной дистрофией роговицы Фукса. Краткий обзор данных 25 , 104047 (2019).
Артикул Google Scholar
Chu, Y. et al. Анализ предсимптоматической ткани для понимания молекулярных и механистических причин позднего дегенеративного тринуклеотидного повторного заболевания. Nucleic Acids Res. 48 , 6740–6758 (2020).
Артикул Google Scholar
Полсон, Х. Болезни повторного распространения. Handb. Clin. Neurol. 147 , 105–123 (2018).
Артикул Google Scholar
Солиман, А. З., Син, К., Радван, С.Х., Гонг, X. и Мутха, В.В. Корреляция тяжести эндотелиальной дистрофии роговицы Фукса с расширением триплетных повторов в TCF4. JAMA Ophthalmol. 133 , 1386–1391 (2015).
З., Син, К., Радван, С.Х., Гонг, X. и Мутха, В.В. Корреляция тяжести эндотелиальной дистрофии роговицы Фукса с расширением триплетных повторов в TCF4. JAMA Ophthalmol. 133 , 1386–1391 (2015).
Артикул Google Scholar
Rohilla, K. J. & Gagnon, K. T. Биология РНК ассоциированных с заболеванием экспансий микросателлитных повторов. Acta Neuropathol. Commun. https://doi.org/10.1186/s40478-017-0468-y (2017).
Артикул PubMed PubMed Central Google Scholar
Du, J. et al. Токсичность РНК и неправильное сплицевание при распространенном заболевании глаз эндотелиальная дистрофия роговицы Фукса. J. Biol. Chem. 290 , 5979–5990 (2015).
CAS Статья Google Scholar
Ронг, З., Ху, Дж., Кори, Д. Р. и Мутха, В. В. Количественные исследования белков мышечной слепоты и их взаимодействия с фокусами РНК TCF4 подтверждают участие в механизме дистрофии Фукса.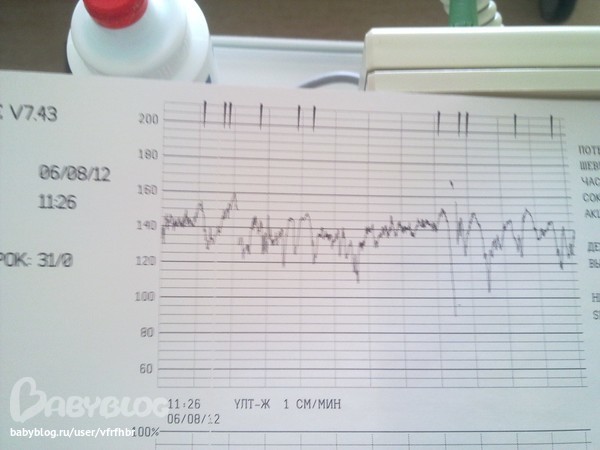 Investig. Офтальмол. Vis. Sci. 60 , 3980–3991 (2019).
Investig. Офтальмол. Vis. Sci. 60 , 3980–3991 (2019).
CAS Статья Google Scholar
Hu, J. et al. Олигонуклеотиды, нацеленные на экспансию триплетных повторов TCF4, ингибируют фокусы РНК и неправильное сплайсинг при дистрофии Фукса. Hum. Мол. Genet. https://doi.org/10.1093/hmg/ddy018 (2018).
Артикул PubMed PubMed Central Google Scholar
Zarouchlioti, C. et al. Антисмысловая терапия распространенной дистрофии роговицы снижает токсичность, опосредованную экспансией повторов TCF4. Am. J. Hum. Genet. https://doi.org/10.1016/j.ajhg.2018.02.010 (2018).
Артикул PubMed PubMed Central Google Scholar
Нельсон, Д. Л., Орр, Х. Т. и Уоррен, С. Т. Нестабильные повторы — три развивающихся лица неврологических заболеваний. Нейрон 77 , 825–843 (2013).
Нейрон 77 , 825–843 (2013).
CAS Статья Google Scholar
Сальседо-Арельяно, М. Дж., Дюфур, Б., МакЛеннан, Ю., Мартинес-Сердено, В. и Хагерман, Р. Синдром ломкой Х-хромосомы и связанные с ним расстройства: Клинические аспекты и патология. Neurobiol. Дис. 136 , 104740 (2020).
CAS Статья Google Scholar
Metsu, S. et al. FRA2A представляет собой экспансию CGG-повтора, связанную с подавлением AFF3. PLoS Genet. 10 , e1004242 (2014).
Артикул Google Scholar
Chutake, Y. K., Lam, C., Costello, W. N., Anderson, M. & Bidichandani, S. I. Подавление эпигенетического промотора при атаксии Фридрейха зависит от длины повтора. Ann. Neurol. 76 , 522–528 (2014).
CAS Статья Google Scholar
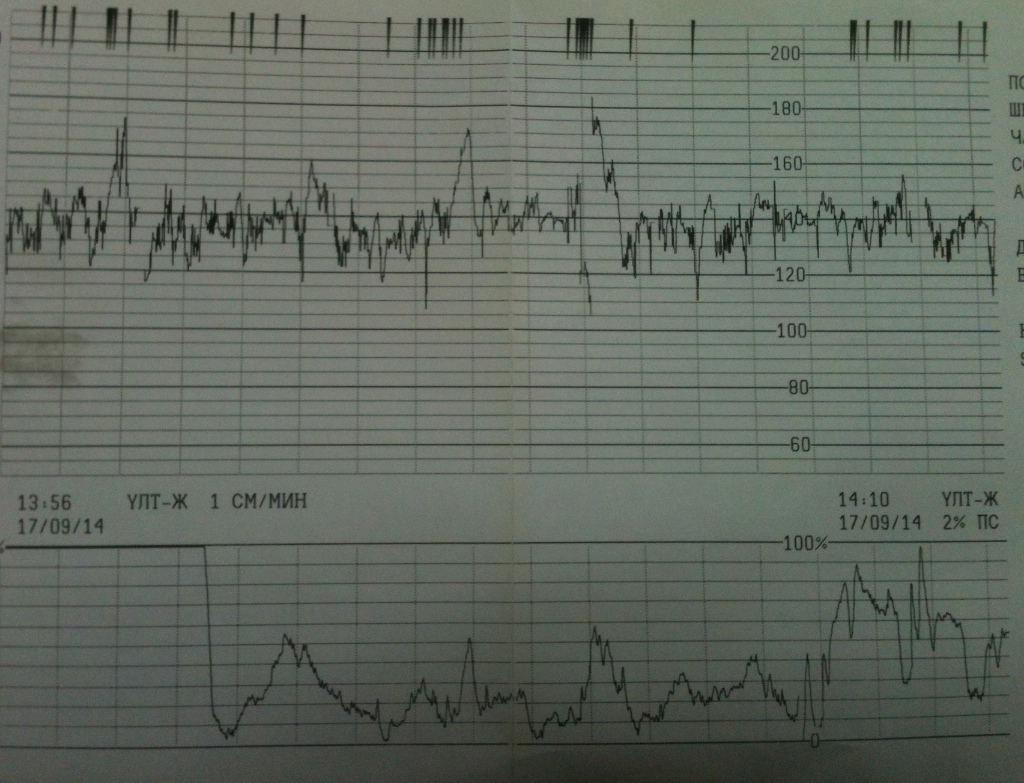
Gijselinck, I. et al. Размер повтора C9orf72 коррелирует с возрастом начала заболевания, метилированием ДНК и подавлением транскрипции промотора. Мол. Психиатрия 21 , 1112–1124 (2016).
CAS Статья Google Scholar
Esvald, E.-E. et al. Факторы транскрипции семейства CREB являются основными медиаторами ауторегуляции транскрипции BDNF в корковых нейронах. J. Neurosci. 40 , 1405–1426 (2020).
CAS Статья Google Scholar
Lizio, M. et al. Пути к атласу экспрессии млекопитающих на уровне промотора FANTOM5. Genome Biol. 16 , 22 (2015).
CAS Статья Google Scholar
Нетранслированная экспансия CTG вызывает новую форму спиноцеребеллярной атаксии (SCA8)
Subramony, S.H. & Currier, R.D.Классификация семейных атаксий.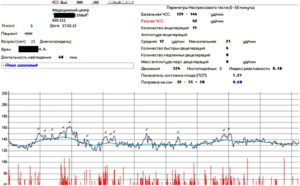 в Справочнике по клинической неврологии (ред. Винкен, П.Дж., Брюн, Г.В. и Клаванс, Х.Л.) 271–284 (Elsevier Science Publishing Company, Нью-Йорк, 1991).
в Справочнике по клинической неврологии (ред. Винкен, П.Дж., Брюн, Г.В. и Клаванс, Х.Л.) 271–284 (Elsevier Science Publishing Company, Нью-Йорк, 1991).
Google Scholar
Harding, A.E. Клинические особенности и классификация аутосомно-доминантной мозжечковой атаксии с поздним началом: исследование 11 семей, включая потомков «семьи Дрю из Уолворта». Мозг 105 , 1-28 (1982).
CAS Статья PubMed Google Scholar
Zoghbi, H.Y. Спиноцеребеллярная дегенерация. Curr. Neurol. 11 , 121–144 (1991).
Google Scholar
Брайс, А. Нестабильные мутации и нейродегенеративные расстройства. J. Neurol. 245 , 505–510 (1998).
CAS Статья PubMed Google Scholar
Клокжедей, Т. и Эверт, Б. Гены, участвующие в наследственных атаксиях. Trends Neurosci. 21 , 413–418 (1998).
и Эверт, Б. Гены, участвующие в наследственных атаксиях. Trends Neurosci. 21 , 413–418 (1998).
CAS Статья PubMed Google Scholar
Jansen, G. et al. Аномальные уровни протеинкиназы миотонической дистрофии вызывают у мышей только легкую миопатию. Nature Genet. 13 , 316–324 (1996).
CAS Статья PubMed Google Scholar
Reddy, S. et al. У мышей, лишенных протеинкиназы миотонической дистрофии, развивается прогрессирующая миопатия с поздним началом. Nature Genet. 13 , 325–335 (1996).
CAS Статья PubMed Google Scholar
Groenen, P. & Wieringa, B.Возрастающая сложность миотонической дистрофии. Bioessays 20 , 901–912 (1998).
CAS Статья PubMed Google Scholar
Hoffmann-Radvanyi, H.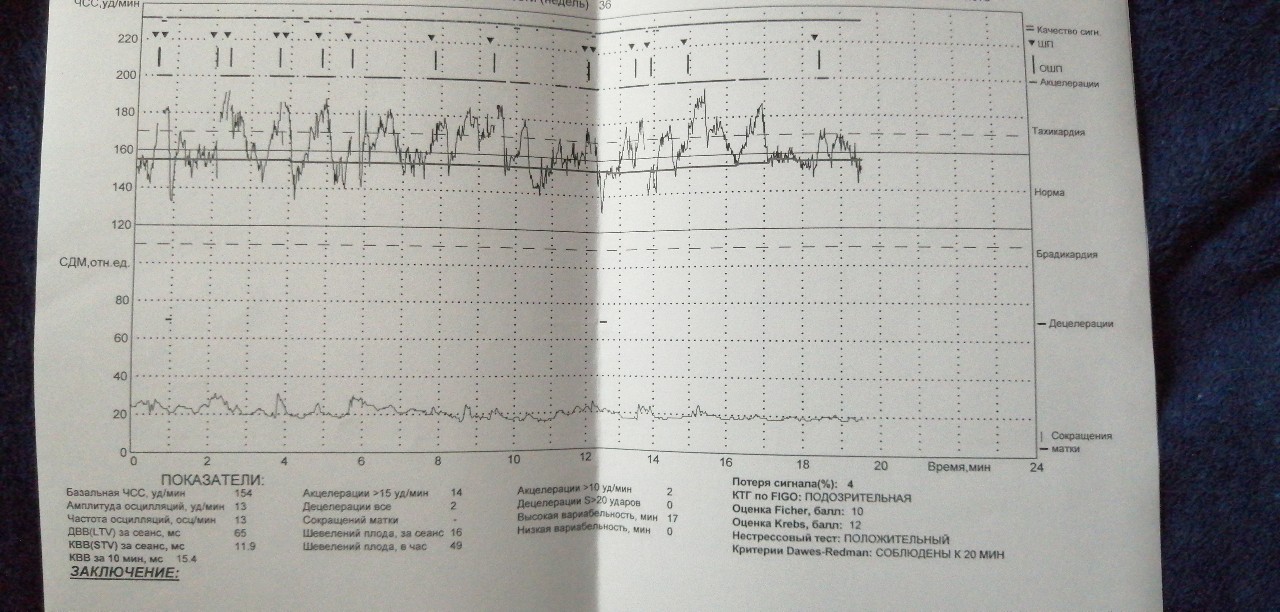 et al. Миотоническая дистрофия: отсутствие увеличенного транскрипта CTG в врожденных формах и низкая экспрессия нормального аллеля. Hum. Мол. Genet. 2 , 1263–1266 (1993).
et al. Миотоническая дистрофия: отсутствие увеличенного транскрипта CTG в врожденных формах и низкая экспрессия нормального аллеля. Hum. Мол. Genet. 2 , 1263–1266 (1993).
Артикул Google Scholar
Krahe, R.и другие. Влияние экспансии тринуклеотидного повтора миотонической дистрофии на транскрипцию и процессинг DMPK. Genomics 28 , 1–14 (1995).
CAS Статья PubMed Google Scholar
Taneja, K.L., McCurrach, M., Schalling, M., Housman, D. & Singer, R.H. Очаги транскриптов тринуклеотидных повторов в ядрах клеток и тканей миотонической дистрофии. J. Cell Biol. 128 , 995–1002 (1995).
CAS Статья Google Scholar
Дэвис, Б.М., МакКеррач, М.Е., Танежа, К.Л., Сингер, Р.Х. и Хаусман, Д.Э. Экспансия тринуклеотидного повтора CUG в 3´ нетранслируемой области транскриптов протеинкиназы миотонической дистрофии приводит к сохранению транскриптов в ядре.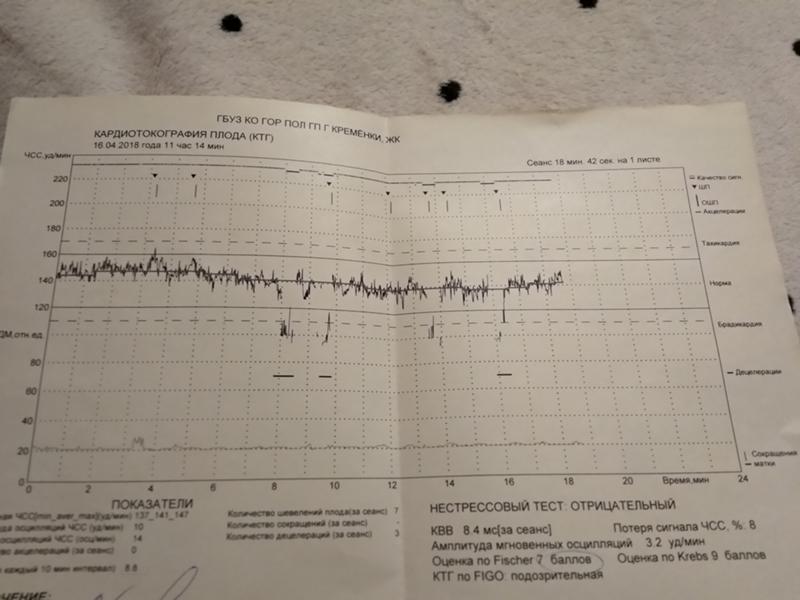 Proc. Natl Acad. Sci. USA 94 , 7388–7393 (1997).
Proc. Natl Acad. Sci. USA 94 , 7388–7393 (1997).
CAS Статья Google Scholar
Тимченко, Л.T. et al. Идентификация РНК-связывающего белка с триплетным повтором (CUG) n и его экспрессия при миотонической дистрофии. Nucleic Acids Res. 24 , 4407–4414 (1996).
CAS Статья PubMed PubMed Central Google Scholar
Roberts, R. et al. Измененное фосфорилирование и внутриклеточное распределение (CUG) n-триплетного повторения РНК-связывающего белка у пациентов с миотонической дистрофией и у мышей с нокаутом миотониновой протеинкиназы. Proc. Natl Acad. Sci. США 94 , 13221–13226 (1997).
CAS Статья PubMed Google Scholar
Филипс, А.В., Тимченко, Л.Т. И Купер, Т. Нарушение сплайсинга, регулируемого CUG-связывающим белком, при миотонической дистрофии. Наука 280 , 737–741 (1998).
Наука 280 , 737–741 (1998).
CAS Статья PubMed PubMed Central Google Scholar
Шаллинг, М., Hudson, T., Buetow, K. & Housman, D. Прямое обнаружение новых увеличенных тринуклеотидных повторов в геноме человека. Nature Genet. 4 , 135–139 (1993).
CAS Статья PubMed Google Scholar
Koob, M.D. et al. Быстрое клонирование расширенных последовательностей тринуклеотидных повторов из геномной ДНК. Nature Genet. 18 , 72–75 (1998).
CAS Статья PubMed Google Scholar
Мозли, М.L. et al. Распространенность доминантных спиноцеребеллярных и триплетных повторов Фридрейха повторяется среди 361 семьи с атаксией. Неврология 51 , 1666–1671 (1998).
CAS Статья PubMed PubMed Central Google Scholar
Chung, M. -y. и другие. Доказательства механизма, предрасполагающего к нестабильности CAG-повторов из поколения в поколение при спиноцеребеллярной атаксии 1 типа. Nature Genet. 5 , 254–258 (1993).
-y. и другие. Доказательства механизма, предрасполагающего к нестабильности CAG-повторов из поколения в поколение при спиноцеребеллярной атаксии 1 типа. Nature Genet. 5 , 254–258 (1993).
CAS Статья PubMed Google Scholar
Cancel, G. et al. Молекулярные и клинические корреляции при спиноцеребеллярной атаксии 2 × исследование 32 семей. Hum. Мол. Genet. 6 , 709–715 (1997).
CAS Статья PubMed Google Scholar
Маруяма, Х. и др. Молекулярные особенности CAG-повторов и клинические проявления болезни Мачадо-Джозефа. Hum. Мол. Genet. 4 , 807–812 (1995).
CAS Статья PubMed Google Scholar
Maciel, P. et al. Корреляция между длиной повтора CAG и клиническими особенностями болезни Мачадо-Джозефа. Am. J. Hum. Genet.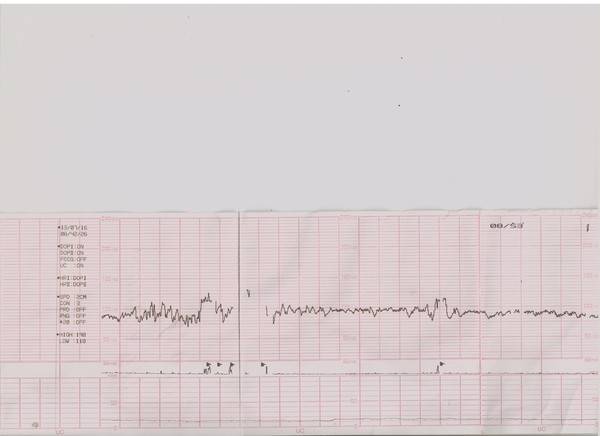 57 , 54–61 (1995).
57 , 54–61 (1995).
CAS PubMed PubMed Central Google Scholar
Жученко, О.и другие. Аутосомно-доминантная мозжечковая атаксия (SCA6), связанная с небольшими разложениями полиглутамина в × -1A-зависимом от напряжения кальциевом канале. Nature Genet. 15 , 62–69 (1997).
CAS Статья PubMed Google Scholar
Jodice, C. et al. Эпизодическая атаксия типа 2 (EA2) и спиноцеребеллярная атксия типа 6 (SCA6) из-за экспансии CAG-повторов в гене CACNA1A на хромосоме 19p. Hum.Мол. Genet. 6 , 1973–1978 (1997).
CAS Статья PubMed Google Scholar
David, G. et al. Молекулярные и клинические корреляции аутосомно-доминантной мозжечковой атаксии с прогрессирующей макулярной дистрофией (SCA7). Hum. Мол. Genet. 7 , 165–170 (1998).
CAS Статья PubMed PubMed Central Google Scholar
Цилфидис, К., MacKenzie, A.E., Mettler, G., Barcelo, J. & Korneluk, R.G. Корреляция между длиной тринуклеотидного повтора CTG и частотой тяжелой врожденной миотонической дистрофии. Nature Genet. 1 , 192–195 (1992).
CAS Статья PubMed Google Scholar
Vanhee-Brossollet, C. & Vaquero, C. Имеют ли смысл естественные антисмысловые транскрипты у эукариот? Gene 211 , 1–9 (1998).
CAS Статья PubMed Google Scholar
Неллен В. и Лихтенштейн К. Что делает мРНК антисмысловой? Trends Biochem. Sci. 18 , 419–423 (1993).
CAS Статья PubMed Google Scholar
Lathrop, G.M., Lalouel, J.M., Julier, C.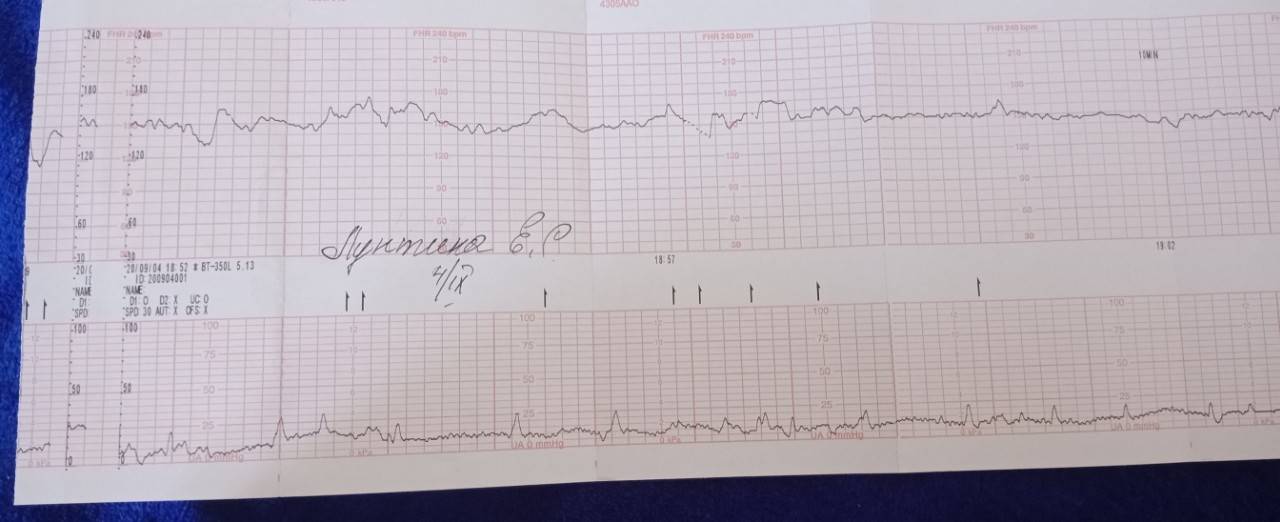 & Ott, J. Стратегии анализа мультилокусных сцеплений у людей. Proc. Natl Acad. Sci. США 81 , 3443–3446 (1984).
& Ott, J. Стратегии анализа мультилокусных сцеплений у людей. Proc. Natl Acad. Sci. США 81 , 3443–3446 (1984).
CAS Статья PubMed Google Scholar
Одна ДНК, два транскрипта, семь рамок считывания, потенциально девять токсичных объектов!
Abstract
Заболевания, связанные с нестабильными повторяющимися элементами в ДНК, РНК и аминокислотах, неизменно преподносят научные сюрпризы. Большинство заболеваний вызвано расширением тринуклеотидных повторов, что в конечном итоге приводит к таким заболеваниям, как болезнь Хантингтона, миотоническая дистрофия, синдром ломкой Х-хромосомы и серия спиноцеребеллярных атаксий.Эти повторяющиеся мутации динамичны, меняются от поколения к поколению и внутри человека, и повторы могут транскрибироваться в двух направлениях. Неожиданные способы патогенеза включают аберрантную потерю экспрессии белка; аберрантная сверхэкспрессия немутантных белков; функция токсического прироста белка через расширенные полиглутаминовые тракты, которые кодируются расширенными трактами CAG; и РНК-токсическое усиление функции, вызванное транскриптами, несущими расширенные участки CUG, CAG или CGG. Недавний прогресс показывает, что транскрипты РНК с расширенными повторами CAG могут транслироваться при полном отсутствии стартового ATG, и эта трансляция, связанная с повторами, не-ATG (RAN-трансляция) происходит через расширенные повторы CAG во всех рамках считывания (CAG, AGC, и GCA) для производства гомополимерных белков длинных полиглутаминовых, полисериновых и полиаланиновых трактов.Расширенные участки CTG, экспрессирующие транскрипты CUG, также показывают RAN-трансляцию, происходящую во всех трех рамках (CUG, UGC и GCU), с образованием полилейцина, полицистеина и полиаланина. Эти продукты RAN-трансляции могут быть токсичными. Таким образом, одна нестабильная (CAG) • (CTG) ДНК может продуцировать два расширенных повторяющихся транскрипта и гомополимерные белки с рамками считывания (AUG-направленный polyGln и шесть белков трансляции RAN), в результате чего получается в общей сложности девять потенциально токсичных объектов. Появление RAN-трансляции в тканях пациента расширяет наши горизонты патогенеза заболеваний.
Недавний прогресс показывает, что транскрипты РНК с расширенными повторами CAG могут транслироваться при полном отсутствии стартового ATG, и эта трансляция, связанная с повторами, не-ATG (RAN-трансляция) происходит через расширенные повторы CAG во всех рамках считывания (CAG, AGC, и GCA) для производства гомополимерных белков длинных полиглутаминовых, полисериновых и полиаланиновых трактов.Расширенные участки CTG, экспрессирующие транскрипты CUG, также показывают RAN-трансляцию, происходящую во всех трех рамках (CUG, UGC и GCU), с образованием полилейцина, полицистеина и полиаланина. Эти продукты RAN-трансляции могут быть токсичными. Таким образом, одна нестабильная (CAG) • (CTG) ДНК может продуцировать два расширенных повторяющихся транскрипта и гомополимерные белки с рамками считывания (AUG-направленный polyGln и шесть белков трансляции RAN), в результате чего получается в общей сложности девять потенциально токсичных объектов. Появление RAN-трансляции в тканях пациента расширяет наши горизонты патогенеза заболеваний.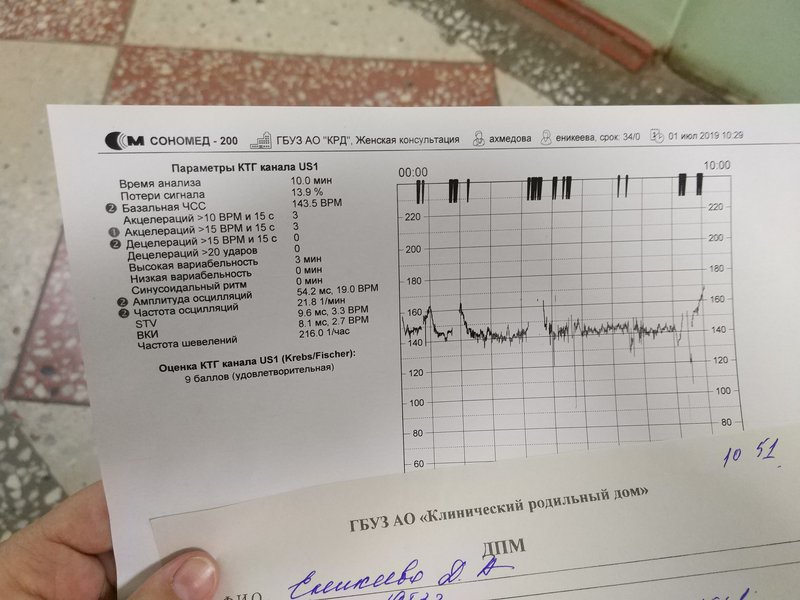 Более того, поскольку RAN-перевод противоречит каноническим требованиям инициации перевода, теперь возникает много новых вопросов, которые необходимо решить. В этом обзоре рассматривается RAN-перевод и некоторые актуальные вопросы.
Более того, поскольку RAN-перевод противоречит каноническим требованиям инициации перевода, теперь возникает много новых вопросов, которые необходимо решить. В этом обзоре рассматривается RAN-перевод и некоторые актуальные вопросы.
Образец цитирования: Pearson CE (2011) Повторное ассоциированное начало трансляции не-ATG: одна ДНК, две транскрипции, семь рамок чтения, потенциально девять токсичных объектов! PLoS Genet 7 (3): e1002018. https://doi.org/10.1371/journal.pgen.1002018
Редактор: Грегори П. Копенгейвер, Университет Северной Каролины в Чапел-Хилл, Соединенные Штаты Америки
Опубликовано: 10 марта 2011 г.
Авторские права: © 2011 Christopher E. Pearson. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Исследования в лаборатории Пирсона поддерживаются Канадскими институтами исследований в области здравоохранения, Ассоциацией мышечной дистрофии США, Канадской ассоциацией мышечной дистрофии и Совместным исследовательским центром мышечной дистрофии имени Пола Веллстона при Рочестерском университете при поддержке NIH. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Автор заявил, что конкурирующих интересов не существует.
Введение
Изобилие научных сюрпризов связано с заболеваниями, связанными с нестабильными повторяющимися элементами в ДНК, РНК и белках. Эти заболевания вызваны расширением или сокращением тринуклеотидных, тетрануклеотидных, пентануклеотидных, додекануклеотидных и макросателлитных повторов (таблица 1). Этот класс болезней, включающий около 40 заболеваний (полный набор см. В дополнительной таблице 1, представленной в López Castel et al. [1]), позволил выявить несколько уникальных и неожиданных открытий: неожиданности включают мутации, которые являются динамическими — постоянно меняющимися как через поколений и внутри человека (Таблица 1) [1], [2].Также существует множество неожиданных способов патогенеза, в том числе: аберрантная потеря экспрессии белка [3]; аберрантная сверхэкспрессия немутантных белков [4] — [6]; функция токсического прироста белка через расширенные полиглутаминовые тракты, которые кодируются расширенными трактами CAG [3]; и РНК-токсическое усиление функции, вызванное транскриптами, несущими расширенные участки CUG или CGG [7], [8]. Известно даже одно заболевание, вызываемое как токсичным полиглутамином, так и токсической РНК, причем оба они возникают в результате двунаправленной транскрипции через комплементарные цепи повторов CAG и CTG (Таблица 1) [9] — [12].Многие участки генома транскрибируются по обеим цепям, включая большинство повторов, связанных с локусами болезни, что позволяет предположить, что множественные токсические сущности (polyGln и токсичные CUG-РНК) могут вносить вклад в патогенез многих заболеваний [9], [12].
В дополнительной таблице 1, представленной в López Castel et al. [1]), позволил выявить несколько уникальных и неожиданных открытий: неожиданности включают мутации, которые являются динамическими — постоянно меняющимися как через поколений и внутри человека (Таблица 1) [1], [2].Также существует множество неожиданных способов патогенеза, в том числе: аберрантная потеря экспрессии белка [3]; аберрантная сверхэкспрессия немутантных белков [4] — [6]; функция токсического прироста белка через расширенные полиглутаминовые тракты, которые кодируются расширенными трактами CAG [3]; и РНК-токсическое усиление функции, вызванное транскриптами, несущими расширенные участки CUG или CGG [7], [8]. Известно даже одно заболевание, вызываемое как токсичным полиглутамином, так и токсической РНК, причем оба они возникают в результате двунаправленной транскрипции через комплементарные цепи повторов CAG и CTG (Таблица 1) [9] — [12].Многие участки генома транскрибируются по обеим цепям, включая большинство повторов, связанных с локусами болезни, что позволяет предположить, что множественные токсические сущности (polyGln и токсичные CUG-РНК) могут вносить вклад в патогенез многих заболеваний [9], [12].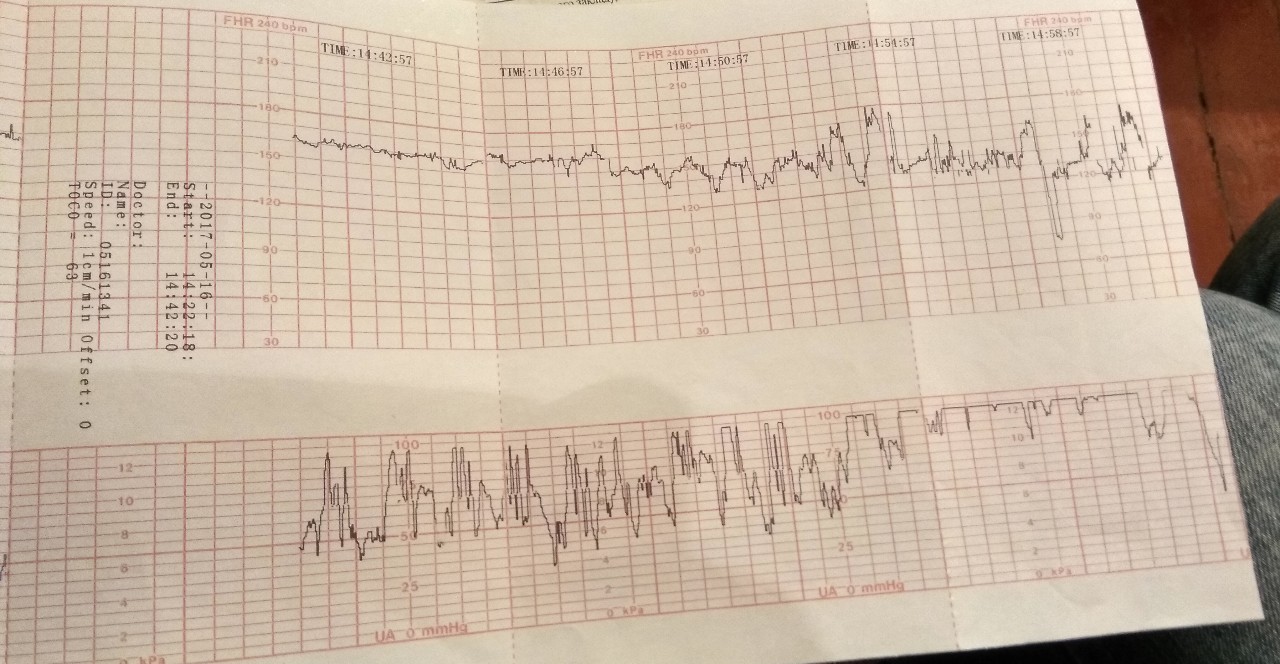 Сколько сюрпризов осталось раскрыть этому полю?
Сколько сюрпризов осталось раскрыть этому полю?
Открытие трансляции не-ATG, связанной с повторами (RAN-трансляция)
Лаборатория доктора Ранума только что опубликовала статью, которая теперь раскрывает еще один сюрприз: транскрипты РНК с расширенными повторами CAG могут транслироваться при полном отсутствии исходного ATG, и эта трансляция, связанная с повторами, не-ATG (RAN-трансляция) происходит через расширенные повторы CAG во всех рамках считывания (CAG, AGC и GCA) с образованием гомополимерных белков длинных полиглутаминовых, полисериновых и полиаланиновых трактов [13] (рис. 1).Конструкции с расширенными трактами CTG, экспрессирующими транскрипты CUG, также показывают RAN-трансляцию, которая также происходит во всех трех кадрах (CUG, UGC и GCU), с образованием полилейцина, полицистеина (рис. 1) и полиаланина. Это противоречит каноническим требованиям инициации перевода [14]. RAN-трансляция происходит на экспансионных конструкциях, которые интегрированы в геном в клетках и мозге, а также в тканях трансгенных мышей на моделях повторяющихся заболеваний. Важно отметить, что Ranum и др. Показывают, что RAN-трансляция происходит в релевантных для заболевания тканях пациентов с SCA8 и DM1 с расширениями CAG / CTG.
Важно отметить, что Ranum и др. Показывают, что RAN-трансляция происходит в релевантных для заболевания тканях пациентов с SCA8 и DM1 с расширениями CAG / CTG.
Шаблоны и ткани для RAN-трансляции
RAN-трансляция зависит от длины повтора в разной степени для каждого кадра считывания. Конструкции, содержащие 15-20 повторов CAG, не экспрессируют polyGln посредством RAN-трансляции, в отличие от конструкций с 42-107 повторами. PolyAla устойчиво экспрессировалась с ~ 100 повторами, умеренно с ~ 80 повторами, но не с 42 и 58 повторами. PolySer был обнаружен с 58–107, но не с 42 повторами. Зависимость от длины для конструкций CTG для RAN-трансляции полилейцина, полицистеина и полиаланина еще предстоит определить.Таким образом, RAN-трансляция зависит от длины, и для данной длины тракта CAG polyAla экспрессируется на самых высоких уровнях, за ней следуют polyGln и polySer. Более длинные участки повторов могут экспрессировать несколько гомополимерных белков, транслируемых RAN. Ранум и его коллеги ясно продемонстрировали, что все три RAN-транслируемых белка могут экспрессироваться одновременно в одной клетке.
Ранум и его коллеги ясно продемонстрировали, что все три RAN-транслируемых белка могут экспрессироваться одновременно в одной клетке.
RAN-трансляция, как было показано, происходит в контексте других релевантных для заболевания последовательностей в виде конструкций с фланкирующими последовательностями выше CAG-повтора из Хантингтина (HD), Хантингтин-подобного 2 (HDL2), спиноцеребеллярной атаксии 3 типа (SCA3), или локусы миотонической дистрофии типа 1 (DM1) показали устойчивую экспрессию polyGln и polyAla и вариабельную экспрессию polySer в отсутствие стартового кодона ATG.
Неожиданное наблюдение RAN-трансляции на участках CAG и CUG может также поставить под вопрос, какие другие повторяющиеся последовательности могут быть разрешающими для RAN-трансляции. Например, могут ли повторы CGG или CCG быть RAN-переведены? FXTAS, где транскрибируются премутации CGG-разложения на FMR1 и антисмысловые тракты CCG от FMR4 (Таблица 1). Пока что данные подтверждают роль режима токсической РНК в патогенезе FXTAS [8]. RAN-трансляция через тракты CGG и CCG представит пробеги polyArg, polyAla, polyGly, polyPro, polyArg и polyAla как потенциально токсичные сущности для FXTAS.
RAN-трансляция через тракты CGG и CCG представит пробеги polyArg, polyAla, polyGly, polyPro, polyArg и polyAla как потенциально токсичные сущности для FXTAS.
Расширение трактов полиаланина и полиаспарагиновой кислоты может вызывать заболевание. По крайней мере девять заболеваний вызываются расширением трактов поли-Ala в белках, а одно вызвано расширением потока полиаспарагиновой кислоты (таблица 1; см. Также [1], [15]). Была выдвинута гипотеза, что причиной болезни является нестабильность других аминокислотных последовательностей [16]. В случаях, когда известен патогенный путь, очевидны как потеря функции, так и усиление функции, и пути доминирования отрицательного белка.
Могут ли более длинные повторяющиеся единицы, такие как расширения тетрануклеотидов или пентануклеотидов, быть склонными к RAN-трансляции? Возможность того, что CUG присутствует в расширенном тракте CCUG транскриптов DM2, может приводить к патогенным RAN-транслируемым продуктам, заслуживает рассмотрения.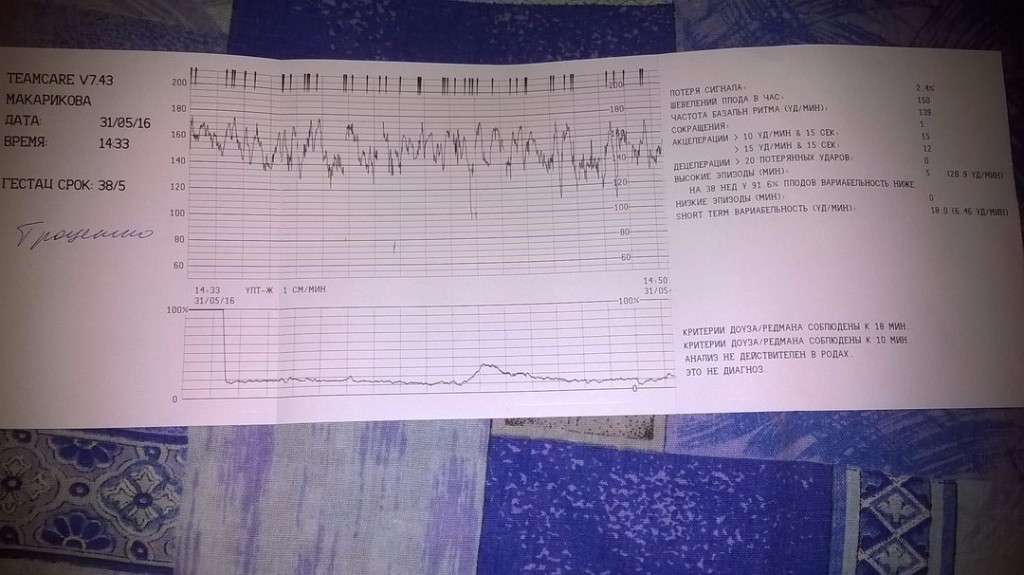 RAN-трансляция через тракты CCUG или CAGG будет давать набор полимерных тетрапептидов, где каждая из четырех возможных рамок считывания будет давать отдельную аминокислоту. CCUG при RAN-трансляции будет производить CCU, GCC, UGC и CUG, создавая повторяющиеся единицы тетрапептида (ProAlaCysLeu) n. CAGG, когда RAN-транслируется, будет продуцировать CAG, GCA, GGC и AGG, создавая повторяющиеся единицы тетрапептида (GlnAlaGlyArg) n. Примечательно, что расширение повторяющихся аминокислотных трактов, как и в случае мутаций инсерционного расширения октапептидного повтора прионного белка (таблица 1), напрямую связано с прионной болезнью: нормальный PrP имеет четыре октапептидных сегмента, тогда как расширенные формы могут иметь столько же в виде девяти дополнительных октапептидных единиц.Генетическое ожидание очевидно в том, что начало прионной болезни коррелирует с размером экспансии октапептида; при наличии до четырех дополнительных единиц возраст дебюта> 60 лет; при пяти-девяти дополнительных единицах возраст дебюта составляет 30-40 лет [17].
RAN-трансляция через тракты CCUG или CAGG будет давать набор полимерных тетрапептидов, где каждая из четырех возможных рамок считывания будет давать отдельную аминокислоту. CCUG при RAN-трансляции будет производить CCU, GCC, UGC и CUG, создавая повторяющиеся единицы тетрапептида (ProAlaCysLeu) n. CAGG, когда RAN-транслируется, будет продуцировать CAG, GCA, GGC и AGG, создавая повторяющиеся единицы тетрапептида (GlnAlaGlyArg) n. Примечательно, что расширение повторяющихся аминокислотных трактов, как и в случае мутаций инсерционного расширения октапептидного повтора прионного белка (таблица 1), напрямую связано с прионной болезнью: нормальный PrP имеет четыре октапептидных сегмента, тогда как расширенные формы могут иметь столько же в виде девяти дополнительных октапептидных единиц.Генетическое ожидание очевидно в том, что начало прионной болезни коррелирует с размером экспансии октапептида; при наличии до четырех дополнительных единиц возраст дебюта> 60 лет; при пяти-девяти дополнительных единицах возраст дебюта составляет 30-40 лет [17].
Продукты RAN-Translation
Каковы функции тандемных последовательностей аминокислот и как они могут иметь отношение к болезни? Многие белки позвоночных содержат тандемные последовательности аминокислот, подавляющее большинство из которых присутствует в факторах метаболизма ДНК / РНК, где, как полагают, гомополимеры участвуют в образовании больших мультибелковых комплексов [18].Такие функции могут соответствовать способности белков polyGln и polyAla образовывать агрегаты и могут указывать на патогенную роль, подобную аберрации в транскриптоме или сплайсингу, как и в случае других повторяющихся заболеваний [3].
В тканях и клетках, специфичных для заболевания, были обнаружены гомополимерные белки, транслируемые с помощью RAN. Антитела, разработанные специально для предсказанных аминокислотных последовательностей ниже транслируемых RAN белков SCA8-polyAla и DM1 polyGln. SCA8-polyAla был обнаружен в соме клеток Пуркинье и дендритах по всему мозжечку на мышиной модели SCA8 и в клетках Пуркинье мозжечка в посмертных образцах от пациентов с SCA8.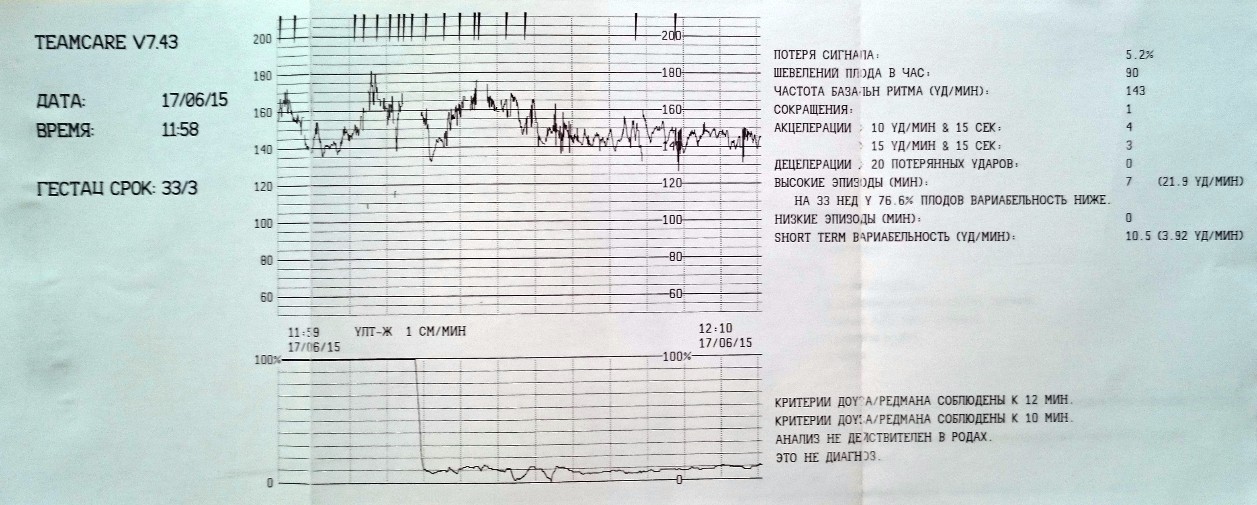 Точно так же в тканях мышей с СД1 и сердечных миоцитах, лейкоцитах и мибластах пациентов с СД1 были обнаружены ядерные агрегаты polyGln. Они не были обнаружены в тканях здоровых или контрольных мышей. Все эти наблюдения подтверждают наличие RAN-транслируемых белков in vivo и в тканях пациента.
Точно так же в тканях мышей с СД1 и сердечных миоцитах, лейкоцитах и мибластах пациентов с СД1 были обнаружены ядерные агрегаты polyGln. Они не были обнаружены в тканях здоровых или контрольных мышей. Все эти наблюдения подтверждают наличие RAN-транслируемых белков in vivo и в тканях пациента.
Продукты трансляции RAN могут быть токсичными и могут быть связаны с патологией заболевания. Zu et al. показали, что клетки, трансфицированные конструкциями CAG, которые транслируются с помощью RAN в polyGln, polyAla и polySer, демонстрируют некоторые признаки повышенного апоптоза, предполагая, что продукты RAN-трансляции могут быть токсичными для клеток и могут способствовать симптомам нейродегенеративного заболевания.Однако из-за использованного исхода токсичности (CAA по сравнению с повторами CAG в качестве меры RAN-трансляции) токсичность потенциально осложняется любой РНК-опосредованной токсичностью. Примечательно, что уже было продемонстрировано, что polyAla токсичен для клеток и может способствовать развитию ряда заболеваний (таблица 1; см. Также [15]). В случае RAN-трансляции неясно, является ли polyAla, polySer или PolyGln причиной явной токсичности и в какой степени это может способствовать заболеванию.
Также [15]). В случае RAN-трансляции неясно, является ли polyAla, polySer или PolyGln причиной явной токсичности и в какой степени это может способствовать заболеванию.
Механизм RAN-трансляции: инициация, РНК-структура,
цис, -элементы и trans -Факторы Механизм инициации RAN-трансляции неясен.Транскрипты CAG без AUG осаждались совместно с легкими полирибосомами. RAN-трансляция была чувствительна к циклогексимиду, который связывается с рибосомами и ингибирует удлинение трансляции. Было бы интересно изучить влияние других соединений (таких как лактимидомицин или эметин), которые могут по-разному влиять на инициацию или удлинение трансляции [19], [20] на RAN-трансляцию. RAN-трансляция отличается от трансляционного сдвига рамки, о котором ранее сообщалось для трактов CAG [21], [22]. Неизвестно, где именно в тракте CAG происходит инициация RAN-трансляции.Сайты инициации RAN-трансляции могут различаться для повторов и рамок считывания. Например, RAN-трансляция каркаса CAG давала преимущественно четкую полосу полиглутаминового белка с молекулярной массой, аналогичной молекулярной массе белка, инициированного стартовым кодоном AUG непосредственно перед повтором [13], что позволяет предположить, что RAN-трансляция инициировалась только при начало повторяющейся последовательности — либо в первом CAG, либо в кодоне, отличном от AUG, непосредственно перед повтором.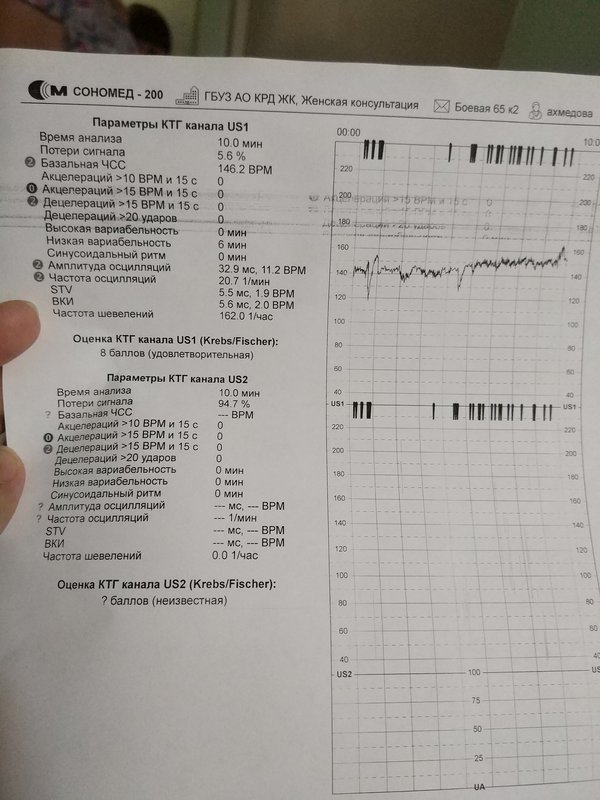 Напротив, RAN-трансляция каркаса GCA дала серию белков с плотными интервалами, что позволяет предположить, что инициация происходила в различных точках повтора.Это было подтверждено масс-спектрометрическим анализом продуктов polyAla RAN, который выявил ряд N-концевых пептидов с различным числом остатков аланина и C-концевой фрагмент с предсказанными C-концевыми остатками. Таким образом, инициация или RAN-трансляция может варьироваться между повторяющимися последовательностями.
Напротив, RAN-трансляция каркаса GCA дала серию белков с плотными интервалами, что позволяет предположить, что инициация происходила в различных точках повтора.Это было подтверждено масс-спектрометрическим анализом продуктов polyAla RAN, который выявил ряд N-концевых пептидов с различным числом остатков аланина и C-концевой фрагмент с предсказанными C-концевыми остатками. Таким образом, инициация или RAN-трансляция может варьироваться между повторяющимися последовательностями.
, вероятно, имеет решающее значение для RAN-трансляции. Об инициации не-AUG сообщалось в небольшом количестве генов млекопитающих, возникающих в различных кодонах, включая ACG, CUG, GUG и UUG [23].Во всех случаях это результат спаривания оснований тРНК метионина с кодоном, дополнительным только по двум основаниям. Напротив, Zu et al. показали, что трансляция RAN, по-видимому, не инициируется через N-концевой метионин [13]. Инициатор Met-tRNA-независимая форма инициации трансляции, отличная от AUG, используется некоторыми вирусами [24]. Эти вирусы используют внутренний сайт входа в рибосомы (IRES), который рекрутирует рибосомы и инициирует трансляцию на сайтах, не относящихся к AUG. Для инициации трансляции необходимо поддержание спаривания оснований в структурах «стебель-петля» псевдоузла IRES.По-видимому, IRES структурно имитирует тРНК инициатора и манипулирует рибосомой, чтобы обеспечить инициацию трансляции, не являющуюся AUG. Будущие исследования покажут, может ли способность расширенных транскриптов CAG предполагать как внутрицепочечные шпильки, так и структуры разветвленной РНК [25], может быть связана с этим вирусным способом инициации не-AUG Met-tRNA-независимой трансляции [24]. Например, длинные r (CAG) тракты могут допускать сворачивание в тРНК-подобные структуры, которые допускают самопрайминг трансляции.Основываясь на зависимости от длины тракта CNG, Ранум и его коллеги выдвигают гипотезу, что инициация RAN-трансляции может включать шпильки РНК [13], поскольку длинные участки повторов CAG и CUG могут образовывать шпильки и разветвленные структуры РНК [25].
Эти вирусы используют внутренний сайт входа в рибосомы (IRES), который рекрутирует рибосомы и инициирует трансляцию на сайтах, не относящихся к AUG. Для инициации трансляции необходимо поддержание спаривания оснований в структурах «стебель-петля» псевдоузла IRES.По-видимому, IRES структурно имитирует тРНК инициатора и манипулирует рибосомой, чтобы обеспечить инициацию трансляции, не являющуюся AUG. Будущие исследования покажут, может ли способность расширенных транскриптов CAG предполагать как внутрицепочечные шпильки, так и структуры разветвленной РНК [25], может быть связана с этим вирусным способом инициации не-AUG Met-tRNA-независимой трансляции [24]. Например, длинные r (CAG) тракты могут допускать сворачивание в тРНК-подобные структуры, которые допускают самопрайминг трансляции.Основываясь на зависимости от длины тракта CNG, Ранум и его коллеги выдвигают гипотезу, что инициация RAN-трансляции может включать шпильки РНК [13], поскольку длинные участки повторов CAG и CUG могут образовывать шпильки и разветвленные структуры РНК [25].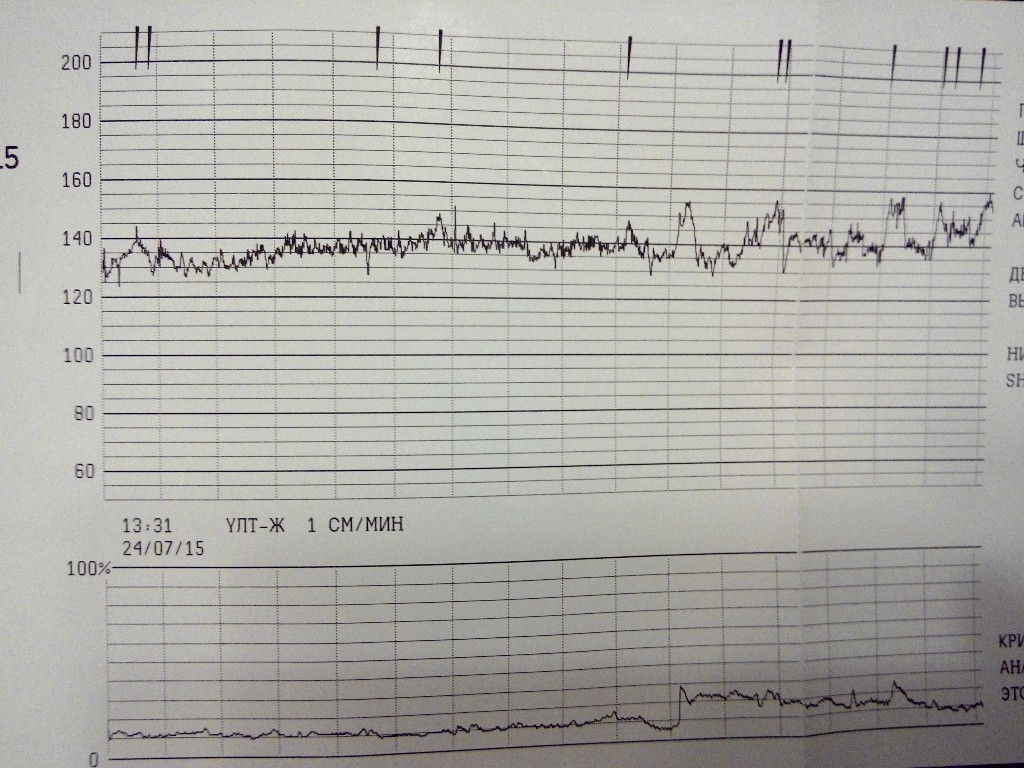 могут экспрессировать гомополимеры в отсутствие повторяющихся последовательностей, начинающихся с ATG, в то время как повторяющиеся последовательности, которые не могут образовывать шпильки, не отображают RAN-трансляцию. Дальнейшее подтверждение роли шпильки CAG / CUG происходит из отчета о том, что шпилька, ранее использовавшаяся в качестве блока трансляции, могла в некоторых контекстах транскрипта FMR1 , лишенного тракта CGG, приводить к инициации трансляции в кодонах GUG в стержень шпильки [26].Подобно остановке трансляции шпилькой Козака [27], сообщалось, что экспансия либо повторов CGG, либо повторов CTG (но не повторов CAG) подавляла трансляцию зависимым от длины тракта и контекста (5’UTR) способом [ 28], [29]. Примечательно, что обнаружение контекстно-зависимой инициации в шпильке Kozak [26], связанной со способностью образовывать шпильки повторов CAG и CUG, подтверждает возможную роль шпильки в RAN-трансляции. Могут ли неповторяющиеся прерывания расширенных повторов CNG [25], [30] — [32], которые могут нарушить внутрицепочечные конформации РНК [25], повлиять на трансляцию RAN? Какие еще второстепенные структуры допускают RAN-перевод? Любопытно, что в отсутствие, но не в присутствии AUG, было предварительно высказано предположение, что кодон CUG ниже расширенного тракта CUG в рамке считывания может инициировать низкие уровни трансляции; однако этого не последовало [29].
могут экспрессировать гомополимеры в отсутствие повторяющихся последовательностей, начинающихся с ATG, в то время как повторяющиеся последовательности, которые не могут образовывать шпильки, не отображают RAN-трансляцию. Дальнейшее подтверждение роли шпильки CAG / CUG происходит из отчета о том, что шпилька, ранее использовавшаяся в качестве блока трансляции, могла в некоторых контекстах транскрипта FMR1 , лишенного тракта CGG, приводить к инициации трансляции в кодонах GUG в стержень шпильки [26].Подобно остановке трансляции шпилькой Козака [27], сообщалось, что экспансия либо повторов CGG, либо повторов CTG (но не повторов CAG) подавляла трансляцию зависимым от длины тракта и контекста (5’UTR) способом [ 28], [29]. Примечательно, что обнаружение контекстно-зависимой инициации в шпильке Kozak [26], связанной со способностью образовывать шпильки повторов CAG и CUG, подтверждает возможную роль шпильки в RAN-трансляции. Могут ли неповторяющиеся прерывания расширенных повторов CNG [25], [30] — [32], которые могут нарушить внутрицепочечные конформации РНК [25], повлиять на трансляцию RAN? Какие еще второстепенные структуры допускают RAN-перевод? Любопытно, что в отсутствие, но не в присутствии AUG, было предварительно высказано предположение, что кодон CUG ниже расширенного тракта CUG в рамке считывания может инициировать низкие уровни трансляции; однако этого не последовало [29]. Точные структурные особенности РНК, участвующие в трансляции RAN, должны быть выяснены.
Точные структурные особенности РНК, участвующие в трансляции RAN, должны быть выяснены.
Контекст повтора может повлиять на RAN-трансляцию. Последовательность фланкирующей РНК может влиять на способность или эффективность RAN-трансляции. Например, Zu et al. [13] обнаружили, что RAN-трансляция мозга в полиглутаминовом каркасе не обнаруживалась в контексте HD или SCA3, немного в последовательности DM1 и HDL2 и много в контексте SCA8. Такие эффекты могут быть связаны с различными структурами РНК, принимаемыми одним и тем же трактом CNG в разных генных контекстах [25].Может ли тракт CCG, кодирующий полипролин, непосредственно 3 ‘от тракта HD CAG, или тракт GGC, кодирующий полиглицин, смежный с трактом SBMA CAG [33], [34], модулировать RAN-трансляцию?
Перспективы будущего — их много
Очевидно, канонические правила трансляции не применимы к участкам экспансии CAG • CTG, поскольку в отсутствие кодона ATG увеличенные тринуклеотидные повторы CAG и CTG могут экспрессировать гомополимерные белки экспансии во всех трех кадрах (Рисунок 1). RAN-трансляция может быть тканеспецифическим явлением, во многом похожим на нестабильность повторов, которая может варьироваться на> 5700 повторов между тканями [2], [35].Продукция антител к предсказанным нижестоящим рамкам считывания каждой цепи ДНК расширенных локусов болезни должна показать, возможна ли RAN-трансляция в этих локусах. Необходимы дальнейшие исследования, чтобы выявить истинную патогенную роль любого из продуктов RAN-трансляции. Понимание механизма RAN-трансляции станет областью активных исследований.
RAN-трансляция может быть тканеспецифическим явлением, во многом похожим на нестабильность повторов, которая может варьироваться на> 5700 повторов между тканями [2], [35].Продукция антител к предсказанным нижестоящим рамкам считывания каждой цепи ДНК расширенных локусов болезни должна показать, возможна ли RAN-трансляция в этих локусах. Необходимы дальнейшие исследования, чтобы выявить истинную патогенную роль любого из продуктов RAN-трансляции. Понимание механизма RAN-трансляции станет областью активных исследований.
С открытием RAN-трансляции теперь возникает много новых вопросов. В дополнение к непосредственным вопросам, например: «Способствуют ли белки, транслируемые с помощью RAN, в патологии заболевания?» и «Каков механизм RAN-трансляции?» или «Сколько фенотипов в клеточных и мышиных моделях болезни с тринуклеотидными повторами является результатом РНК-транслируемых белков, а не предполагаемой РНК polyGln или CUG?».К другим вопросам относятся: «Какие еще тринуклеотидные повторы (связанные с заболеванием или иным образом) могут быть RAN-транслированы?» или «Могут ли нетринуклеотидные повторы, такие как тетрануклеотид, пентануклеотид или другие сателлиты, подвергаться RAN-трансляции?» или «Могут ли определенные неповторяющиеся кодоны быть достаточными для инициации RAN-трансляции?» или «Могут ли аминокислотные последовательности ниже RAN-транслируемых гомополимерных трактов способствовать заболеванию?» или «Какие стоп-кодоны используются для продуктов, переведенных в RAN?» или «Могут ли белки, транслируемые с помощью RAN, иметь естественную функцию?» или «какие белки участвуют в RAN-трансляции?» или «Можно ли ожидать истощения пулов аминокислот, если RAN-трансляция превышает емкость клеток, может ли это быть связано с уменьшением аминокислот в плазме крови у пациентов с HD или DM1 и RAN-трансляцией [36], [37]?», и много других.
Выводы
Подтверждение этих первоначальных и захватывающих наблюдений за RAN-трансляцией [13] и их распространение на клиническое заболевание у пациентов сделает эту статью знаменательной, которая изменит большую часть исследований нарушений с повторением нуклеотидов, а также изменит ее форму. сфокусировать наше понимание инициации трансляции и концепции продуктов генов / растяжения ДНК. Открытие RAN-перевода, вероятно, поставит множество лабораторий во многих областях исследований для поиска этих ответов.
Рисунок 1. Запуск RAN-гена.
Одна ДНК, два транскрипта, семь возможных рамок считывания, потенциально девять токсичных объектов! Транскрипты CAG и CTG могут быть токсичными [12], [38], рамка считывания белка polyGln, инициированная AUG, может быть токсичной, и каждая из трех рамок считывания транскрипта CAG или CUG представляет шесть дополнительных гомополимерных белков, что делает всего девять потенциально токсичных сущностей!
https://doi.org/10.1371/journal.pgen.1002018.g001
Благодарности
Я хотел бы поблагодарить моего бывшего профессора, доктора Нахума Зоненберга за содержательные обсуждения.
Ссылки
- 1. López Castel A, Cleary JD, Pearson CE (2010) Повторяющаяся нестабильность как основа для болезней человека и как потенциальная цель для терапии. Nat Rev Mol Cell Biol 11: 165–170.
- 2. López Castel A, Nakamori M, Tomé S, Chitayat D, Gourdon G, Thornton CA, Pearson CE (2010) Расширенный повтор CTG разграничивает границу аномального метилирования CpG в тканях пациентов с миотонической дистрофией.Hum Mol Genet 20: 1–15.
- 3. La Spada AR, Taylor JP (2010) Болезнь повторного распространения: прогресс и загадки в патогенезе болезни. Нат Рев Женет 11: 247–258.
- 4. Pearson CE (2010) FSHD: болезнь повторных сокращений, наконец, готовая к расширению (наше понимание ее патогенеза). PLoS Genet 6: e1001180.
- 5. Леммерс Р.Дж., ван дер Влит П.Дж., Клоостер Р., Саккони С., Каманьо П. и др. (2010) Унифицированная генетическая модель фациоскапуло-плечевой мышечной дистрофии.Наука 329: 1650–1653.
- 6. Снайдер Л., Гэн Л.Н., Леммерс Р.Дж., Киба М., Уэр CB и др. (2010) Facioscapulohumeral dystrophy: неполное подавление ретротранспонированного гена. PLoS Genet 6: e1001181.
- 7. Shin J, Charizanis K, Swanson MS (2009) Патогенные РНК в болезни распространения микросателлитов. Neurosci Lett 466: 99–102.
- 8. Остра Б.А., Виллемсен Р. (2009) FMR1: ген с тремя лицами. Biochim Biophys Acta 1790: 467–477.
- 9. Merienne K, Trottier Y (2009) Расширения SCA8 CAG / CTG, рассказ о двух ТОКСИЧНОСТИ: уникальный или общий случай? PLoS Genet 5: e1000593.
- 10. Дочери Р.С., Таттл Д.Л., Гао В., Икеда Ю., Мозли М.Л. и др. (2009) Увеличение функции РНК при спиноцеребеллярной атаксии 8 типа. PLoS Genet 5: e1000600.
- 11. Moseley ML, Schut LJ, Bird TD, Koob MD, Day JW, Ranum LP (2000) SCA8 CTG repeat: в массе сокращений в сперматозоидах и изменения последовательности между поколениями могут играть роль в снижении пенетрантности.Hum Mol Genet 9: 2125–2130.
- 12. Batra R, Charizanis K, Swanson MS (2010) Партнеры в преступлении: двунаправленная транскрипция при нестабильной микросателлитной болезни. Hum Mol Genet 19 (R1): R77 – R82.
- 13. Зу Т., Гиббенс Б., Доти Н.С., Гомеш-Перейра М., Хугет А. и др. (2010) Не-ATG инициированная трансляция, направляемая расширениями микросателлитов Proc Natl Acad Sci U S. A 108: 260–265.
- 14. Соненберг Н., Хиннебуш А.Г. (2009) Регулирование инициации трансляции у эукариот: механизмы и биологические мишени.Ячейка 136: 731–745.
- 15. Messaed C, Rouleau GA (2009) Молекулярные механизмы, лежащие в основе полиаланиновых заболеваний. Нейробиол Дис. 34: 397–405.
- 16. Karlin S, Burge C (1996) Тринуклеотидные повторы и длинные гомопептиды в генах и белках, связанных с заболеванием и развитием нервной системы. Proc Natl Acad Sci U S A 93: 1560–1565.
- 17. Стивенс Д. Д., Уолтер Э. Д., Родригес А., Дрейпер Д., Дэвис П. и др. (2009) Раннее начало прионной болезни в результате экспансии октопоротов коррелирует со свойствами связывания меди.PLoS Pathog 5: e1000390.
- 18. Faux NG, Bottomley SP, Lesk AM, Irving JA, Morrison JR, de la Banda MG, Whisstock JC (2005) Функциональные сведения о распределении и роли белков, содержащих гомопептидные повторы. Genome Res 15: 537–551.
- 19. Schneider-Poetsch T, Ju J, Eyler DE, Dang Y, Bhat S и др. (2010) Ингибирование элонгации эукариотической трансляции цикогексимидом и лактимидомицином. Nat Chem Bio 6: 209–217.
- 20. Олейник Н.Л. (1977) Инициирование и удлинение синтеза белка в растущих клетках: дифференциальное ингибирование циклогексимидом и эметином.Arch Biochem Biophys 182: 171–80.
- 21. Гаспар С., Джаннатипур М., Дион П., Лаганьер Дж., Секейрос Дж. И др. (2000) CAG-тракт MJD-1 может быть подвержен сдвигам рамки считывания, вызывая накопление полиаланина. Hum Mol Genet 9: 1957–1966.
- 22. Davies JE, Rubinsztein DC (2006) Полиаланин и продукты сдвига рамки полисерина при болезни Хантингтона. J Med Genet 43: 893–896. Ошибка в: J Med Genet 44: 160.
- 23. Touriol C, Bornes S, Bonnal S, Audigier S, Prats H и др.(2003) Создание разнообразия изоформ белка путем альтернативной инициации трансляции в кодонах, не относящихся к AUG. Biol Cell 95: 169–178.
- 24. Ян, E (2006) Дивергентные элементы IRES у беспозвоночных. Исследование вирусов 119: 16–28.
- 25. Sobczak K, Krzyzosiak WJ (2005) CAG-повторы, содержащие прерывания CAA, образуют разветвленные шпильчатые структуры в транскриптах спиноцеребеллярной атаксии типа 2. J Biol Chem 280: 3898–3910.
- 26. Ludwig AL, Hershey JW, Hagerman PJ (2011) Инициация трансляции мРНК FMR1 происходит преимущественно посредством 5′-концевого зависимого сканирования рибосом.J Mol Biol 12 января: Epub выходит в печать. PMID: 21237174.
- 27. Козак М. (1989) Обстоятельства и механизмы ингибирования трансляции вторичной структурой в мРНК эукариот. Mol Cell Biol 9: 5134–5142.
- 28. Feng Y, Zhang F, Lokey LK, Chastain JL, Lakkis L и др. (1995) Подавление трансляции за счет экспансии тринуклеотидных повторов в FMR1. Наука 268: 731–734.
- 29. Raca G, Siyanova EY, McMurray CT, Mirkin SM (2000) Экспансия (CTG) (n) повтора в 5′-UTR репортерного гена препятствует трансляции.Nucleic Acids Res 28: 3943–3949.
- 30. Мозли М.Л., Зу Т., Икеда Й., Гао В., Мосемиллер А.К. и др. (2006) Двунаправленная экспрессия транскриптов экспансии CUG и CAG и внутриядерных включений полиглутамина при спиноцеребеллярной атаксии 8 типа. Nat Genet 38: 758–769.
- 31. Мусова З., Мазанец Р., Крепелова А., Элер Э., Валес Дж. И др. (2009) Очень нестабильные прерывания последовательности CTG-повтора в гене миотонической дистрофии. Am J Med Genet A 149A: 1365–1374.
- 32. Брайда С., Стефанатос Р.К., Адам Б., Махаджан Н., Смитс Х.Дж. и др. (2010) Варианты CCG и GGC повторы в пределах CTG резко изменяют динамику мутаций и, вероятно, способствуют возникновению необычных симптомов у некоторых пациентов с миотонической дистрофией 1 типа. Hum Mol Genet 19: 1399–1412.
- 33. Эндрю С.Е., Голдберг Ю.П., Тейлманн Дж., Цейслер Дж., Хайден М.Р. (1994) Полиморфизм повтора CCG, прилегающий к повтору CAG в гене болезни Хантингтона: последствия для диагностической точности и прогностического тестирования.Hum Mol Genet 3: 65–67.
- 34. Irvine RA, Yu MC, Ross RK, Coetzee GA (1995) микросателлиты CAG и GGC гена рецептора андрогенов находятся в неравновесном сцеплении у мужчин с раком простаты. Cancer Res 55: 1937–1940.
- 35. Клири Дж. Д., Томе С., Лопес Кастель А., Паниграхи Г. Б., Фойри Л. и др. (2010) Тканевые и возрастные паттерны репликации ДНК в локусе CTG / CAG-расширенной миотонической дистрофии человека типа 1. Nat Struct Mol Biol 17: 1079–1087.
- 36. Reilmann R, Rolf LH, Lange HW (1995) Снижение уровня аланина и изолейцина в плазме при болезни Хантингтона. Acta Neurol Scand 91: 222–224.
- 37. Moxley RT 3rd, Kingston W, Griggs RC (1985) Аномальная регуляция венозного аланина после приема глюкозы при миотонической дистрофии. Clin Sci 68: 151–157.
- 38. Li LB, Yu Z, Teng X, Bonini NM (2008) Токсичность РНК является компонентом дегенерации атаксина-3 у Drosophila . Природа 453: 1107–1111.
(PDF) Транскрипция и ядерный транспорт тринуклеотидных повторов CAG / CTG в дрожжах
количество проскальзывания, или, альтернативно, факторы хроматина
по-разному собраны на плазмиде и на хромосоме
, что может приводить к различным свойствам
аппараты транскрипции.
Мы исследовали, подвержены ли естественные тринуклеотидные повторы в
дрожжах проскальзыванию транскрипции.В дрожжах не существует
природных совершенных тринуклеотидных повторов с длиной со- YPL009c содержит участок из
33 несовершенных тринуклеотидных повторов GAA (один из самых длинных
тринуклеотидных повторов в геноме дрожжей), кодирующий либо
,глутаминовой кислоты, либо лизин. МРНК нормального размера, но не L-РНК
, была обнаружена на нозерн-блотах для этого гена (данные не показаны),
, что предполагает, что вырожденные повторы имеют тенденцию уменьшать проскальзывание,
или что существует порог длины, ниже которого проскальзывание не может
или, или повторы GAA менее склонны к проскальзыванию.Эти
трех гипотез не исключают друг друга.
Проскальзывание транскрипции в тринуклеотидных повторах может также происходить в системах млекопитающих.
. Анализ CTG-содержащих
транскриптовв нормальных и миотонических миобластах показал большую вариацию размера
мРНК мутантного DMPK (8). Это было
, что связано с гетерогенностью по размеру повторов на уровне ДНК,
— среди культивированных ®бробластов. Однако возможно, что
некоторых из наблюдаемых молекул мРНК привели к
в результате проскальзывания транскрипции.
В предыдущих исследованиях, в которых авторы изучали клеточное распределение
транскриптов, содержащих длинные тринуклеотидные повторы,
было показано, что, хотя длинные транскрипты
были правильно сплайсированы и полиаденилированы, они не были правильно экспортированы в цитоплазму
. Сообщалось, что мРНК, содержащие CUG,
накапливаются в ядрах культивируемых при DM миобластов (7,8,11)
и в ядрах мышечных клеток трансгенных мышей, экспрессирующих
нетранслируемых повторов CUG (9).Совсем недавно было показано
, что миотоническая дистрофия 2, вызванная экспансией тетрануклеотидного повтора
CCTG в первом интроне гена ZNF9
, также обнаруживает ядерную задержку транскриптов (10). В дрожжах
мы обнаружили, что L-РНК очищаются совместно с фракцией поли (A)
+
, что позволяет предположить, что они полиаденилированы, но мРНК, содержащие CAG и
CUG, не обнаруживают каких-либо явных дефектов в
.ядерный экспорт, хотя они имеют тенденцию образовывать кластеры в клетках
дрожжей, по-видимому, не специфично для какого-либо субклеточного компартмента.
Как CAG-, так и CUG-содержащие транскрипты агрегируются в
дрожжах, тогда как у людей к настоящему времени задокументированы только CUG-содержащие транскрипты
кластеров. Какова природа
этихкластеров? Было высказано предположение, что взаимодействия между
расширенных повторов CUG и двухцепочечной РНК, связывающей
белков, таких как гомологи белка мышечной слепоты дрозофилы
или белка PKR (последний из них имеет гомологи дрожжей), могут объяснить
ядерной задержки мутантных транскриптов DMPK
(11,35).Альтернативно, они могут возникать, когда транскрипты, содержащие тринуклеотид
, экспрессируются из-за межмолекулярных взаимодействий между такими РНК, причем обе гипотезы
не исключают друг друга.
ДОПОЛНИТЕЛЬНЫЙ МАТЕРИАЛ
Дополнительные материалы доступны в NAR Online.
БЛАГОДАРНОСТИ
Мы благодарим J. Boyer за щедрый дар плазмиды
pCMha191, M. Richetti и G. Fischer за критическое прочтение
рукописи и наших коллег за плодотворные обсуждения.
Мы также благодарим Б. Аркангиоли и А. Жакье за соответствующие технические советы
и вдумчивые комментарии к настоящей работе
. B.D. является членом Institut Universitaire de France.
СПРАВОЧНАЯ ИНФОРМАЦИЯ
1. Каммингс, К.Дж. и Зогби, Х. (2000) Четырнадцать и подсчет:
раскрывающих тринуклеотидных повторных заболевания. Гм. Мол. Genet., 9, 909 ± 916.
2. Альба М.М., Сантибанес-Кореф М.Ф. и Хэнкок, Дж. (1999) Повторы аминокислот
в дрожжах чрезмерно представлены в определенных классах белков
и демонстрируют свидетельства мутационного процесса, подобного проскальзыванию.J. Mol.
Эволюция, 49, 789 ± 797.
3. Брук, Дж. Д., МакКеррач, М. Э., Харли, Х. Дж., Баклер, А. Дж., Черч, Д.,
Абуратани, Х., Хантер, К., Стэнтон, В. П., Тирион, Дж. П., Хадсон, Т. и другие.
(1992) Молекулярная основа миотонической дистрофии: экспансия тринуклеотидного повтора
(CTG) на 3′-конце транскрипта, кодирующего белок
, член семейства киназ. Ячейка, 68, 799 ± 808.
4. Харли, Х. Дж., Брук, Дж. Д., Рандл, С. А., Кроу, С., Рирдон, В.,
Баклер, А.Дж., Харпер П.С., Хаусман Д. и Шоу, Д.Дж. (1992)
Расширение нестабильной области ДНК и фенотипические вариации при миотонической дистрофии
. Природа, 355, 545 ± 546.
5. Куб, М. Д., Мозли, М. Л., Шут, Л. Дж., Бензоу, К. А., Берд, Т. Д.,
Дэй, Дж. У. и Ранум, L.P.W. (1999) Нетранслированное расширение CTG
вызывает новую форму спиноцеребеллярной атаксии (SCA8). Nature Genet., 21,
379 ± 384.
6. Krahe, R., Ashizawa, T., Abbruzzese, C., Roeder, E., Carango, P.,
Giacanelli, M., Funanage, V.L. и Сицилиано М.Дж. (1995) Эффект экспансии тринуклеотидного повтора миотонической дистрофии
на транскрипцию и процессинг DMPK
. Геномика, 28, 1-14.
7. Танеха, К.Л., МакКеррач, М., Шаллинг, М., Хаусман, Д. и Сингер Р.
(1995) Очаги транскриптов тринуклеотидных повторов в ядрах миотонических клеток и тканей
дистрофии. J. Cell Biol., 128, 995 ± 1002.
8. Дэвис Б.М., МакКеррач М.Э., Танежа К.Л., Сингер Р.Х. и
Housman, D.E. (1997) Экспансия тринуклеотидного повтора CUG в
3 нетранслируемой области транскриптов протеинкиназы миотонической дистрофии
приводит к сохранению транскриптов в ядре. Proc. Natl Acad. Sci. США, 94,
7388 ± 7393.
9. Манкоди, А., Лоджиан, Э., Каллахан, Л., Макклейн, К., Уайт, Р.,
Хендерсон, Д., Крым, М. и Торнтон, К.А. (2000) Миотоническая дистрофия
у трансгенных мышей, экспрессирующих увеличенный повтор CUG.Наука, 289,
1769 ± 1772.
10. Ликури, К.Л., Рикер, К., Мозли, М.Л., Якобсен, Дж.Ф., Кресс, В.,
Нейлор, С.Л., Дэй, Дж. У. и Ранум, L.P.W. (2001) Миотоническая дистрофия
типа 2, вызванная экспансией CCTG в интроне 1 ZNF9. Наука, 293,
864 ± 867.
11. Миллер, Дж. У., Урбинати, С. Р., Тенг-умнуай, П., Стенберг, М. Г., Бирн, Б. Дж.,
Торнтон, К. и Swanson, M.S. (2000) Рекрутмент человеческих белков, связывающих мышцы
, в экспансии (CUG) n, связанные с миотонической дистрофией
.EMBO J., 19, 4439 ± 4448.
12. Парсонс, М.А., Синден, Р.Р. и Избань М. (1998) Транскрипционные свойства
РНК-полимеразы II в ДНК, содержащей триплетные повторы,
из миотонической дистрофии человека и ломких Х-локусов. J. Biol. Chem.,
273, 26998 ± 27008.
13. Макдональд, Дж. Ф. (1993) Эволюция и последствия сменных элементов
. Curr. Opin. Genet. Дев., 3, 855 ± 864.
14. Северинов, К. и Гольдфарб А. (1994) Топология сайта связывания продукта
в РНК-полимеразе, выявленная по проскальзыванию транскрипта на промоторе фага l P
L
.J. Biol. Chem., 269, 31701 ± 31705.
15. Jeong, S.W., Lang, W.H. и Reeder, R.H. (1996) Дрожжевой терминатор транскрипции
для РНК-полимеразы I разработан для предотвращения проскальзывания полимеразы
. J. Biol. Chem., 271, 16104 ± 16110.
16. Дэн, Л. и Шуман, С. (1997) Свойства удлинения вируса осповакцины
РНК-полимераза: пауза, проскальзывание, добавление 3 ¢ конца и сайт терминации
на выбор. Биохимия, 36, 15892 ± 15899.
17. Вагнер, Л.А., Вайс, Р. Б., Дрисколл, Р., Данн, Д.С. и Гестеланд Р.Ф.
(1990) Проскальзывание транскрипции происходит во время элонгации при пробегах
аденина или тимина в Escherichia coli. Nucleic Acids Res., 18,
3529 ± 3535.
18. Xiong, X.F. и Резникофф, В. (1993) Проскальзывание транскрипции во время процесса инициации транскрипции на мутантном промоторе lac in vivo.
J. Mol. Биол., 231, 569 ± 580.
3546 Nucleic Acids Research, 2002, Vol.30 No. 16
Фреймы для перевода и открытого чтения
Перевод и поиск по рамке открытого чтения
участков ДНК, которые кодируют белки, сначала транскрибируются, в информационную РНК, а затем транслируют в белок. Изучая только последовательность ДНК, мы можем определить последовательность аминокислот, которые появятся в конечном белке. При трансляции кодоны из трех нуклеотидов определяют, какая аминокислота будет добавлена следующей в растущую белковую цепь.Затем важно решить, с какого нуклеотида начать трансляцию и когда остановиться. Это называется открытой рамкой считывания .
После секвенирования гена важно определить правильную открытую рамку считывания (ORF). Каждая область ДНК имеет шесть возможных рамок считывания , по три в каждом направлении. Используемая рамка считывания определяет, какие аминокислоты будут кодироваться геном. Обычно при трансляции гена используется только одна рамка считывания (у эукариот), и часто это самая длинная открытая рамка считывания.Как только открытая рамка считывания известна, последовательность ДНК можно транслировать в соответствующую аминокислотную последовательность. Открытая рамка считывания начинается с atg (Met) у большинства видов и заканчивается стоп-кодоном (taa, tag или tga).
Например, следующая последовательность ДНК может быть прочитана в шести рамках считывания. Три в прямом и три в обратном направлении. Три рамки считывания в прямом направлении показаны с транслированными аминокислотами под каждой последовательностью ДНК.Рамка 1 начинается с буквы «a», рамка 2 начинается с буквы «t», а рамка 3 начинается с буквы «g». Стоп-кодоны обозначены знаком «*» в последовательности белка. Самый длинный ORF находится в кадре 1 .
5 ‘3’
atgcccaagctgaatagcgtagaggggttttcatttgaggacgatgtataa
1 atg ccc aag ctg aat agc gta gag ggg ttt tca tca tca fca ttt gag gac 9 N17 MTA E ta P GAC S *
2 tgc cca agc tga ata gcg tag agg ggt ttt cat cat ttg agg acg atg tat
C P S * I A * R G F H H L R T M Y
3 gcc caa gct gaa tag cgt aga ggg gtt ttc atc att tga gga cga tgt ata
A Q A E * R R G V F I I * G R C I
Практические задачи и Учебное пособие по переводу находятся в Теоретическом разделе.
Для перевода последовательности ДНК мы используем программу SIXFRAME на Верстак биологии. Или вы можете посетить сайт напрямую http://searchlauncher.bcm.tmc.edu/seq-util/Options/sixframe.html
1. Войдите в Biology Workbench и возобновите или создайте новый сеанс.
2. Выберите Nucleic Tools, а затем выберите последовательность, которую вы хотите перевести, установив флажок рядом с файлом.
3.Выберите SIXFRAME, а затем выберите «Выполнить».
4. Вам будет предоставлено окно с просьбой выбрать различные параметры. По умолчанию транслируются все шесть рамок считывания всей последовательности ДНК. Если вы выберете «Показать самую длинную открытую рамку считывания», программа автоматически выберет самую длинную рамку считывания, начиная со стартового кодона (ATG) и заканчивая стоп-кодоном (TAA, TAG или TGA). Это очень удобно.
5. После выбора параметров нажмите «Отправить».
Интерпретация результатов
| ENST00000396203.7 | RPL14-202 | 7136 | 215aa | ENSP00000379506.2 | Ген / транскрипт, который содержит открытую рамку считывания (ORF). Кодирование белка | CCDS43070 | 699CCDS43070 | 99 918. Соответствующая аннотация от NCBI и EMBL-EBI — это результат сотрудничества Ensembl / GENCODE и RefSeq. MANE Select — это транскрипт по умолчанию для гена человека, который является репрезентативным для биологии, хорошо поддерживается, экспрессируется и высококонсервативен.Этот набор транскриптов соответствует GRCh48 и на 100% идентичен между RefSeq и Ensembl / GENCODE для 5 ‘UTR, CDS, сплайсинга и 3’UTR.MANE Select v0.95, единственный транскрипт, выбранный для гена, который является наиболее консервативным, наиболее высокоэффективным. выражен, имеет самую длинную последовательность кодирования и представлен в других ключевых ресурсах, таких как NCBI и UniProt. Это подробно описано на http://www.ensembl.org/info/genome/genebuild/canonical.htmlEnsembl Canonical, набор GENCODE — это набор генов для человека и мыши. представляет собой подмножество репрезентативных транскриптов (варианты сплайсинга).GENCODE basic, APPRIS P1: Ожидается, что транскрипты будут кодировать основную функциональную изоформу, основанную исключительно на основных модулях в APPRIS. APPRIS — это система для аннотирования транскриптов с альтернативным сплайсингом на основе ряда вычислительных методов для идентификации наиболее функционально важных транскриптов гена. APPRIS P1,TSL 1: транскрипт, в котором все сплайсинговые соединения поддерживаются по крайней мере одной не подозреваемой мРНК. Уровень поддержки стенограммы (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемых моделей стенограммы для пользователей, основанный на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 1, | ||
| ENST00000338970.10 | RPL14-201 | 7072 | 215aa | ENSP00000345156.6 | Ген / транскрипт, содержащий открытую рамку считывания | (ORF) .397Protein. | P50914 | — | Набор GENCODE — это набор генов для человека и мыши. представляет собой подмножество репрезентативных транскриптов (вариантов сплайсинга). GENCODE basic, APPRIS P1: Ожидается, что транскрипты будут кодировать основную функциональную изоформу на основе исключительно основных модулей в APPRIS. APPRIS — это система для аннотирования транскриптов с альтернативным сплайсингом на основе ряда вычислительных методов для идентификации наиболее функционально важных транскриптов гена. APPRIS P1,TSL 1: транскрипт, в котором все сплайсинговые соединения поддерживаются по крайней мере одной не подозреваемой мРНК. Уровень поддержки стенограммы (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемых моделей стенограммы для пользователей, основанный на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 1, | |
| ENST00000416518.1 | RPL14-203 | 812 | 124aa | ENSP000003.1 | Ген / транскрипт, содержащий открытую рамку считывания — белок | (ORF 918). | E7EPB3 | — | Набор GENCODE — это набор генов для человека и мыши. представляет собой подмножество репрезентативных транскриптов (варианты сплайсинга) .GENCODE basic, TSL 3: расшифровка, в которой единственной поддержкой является один EST Уровень поддержки транскриптов (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемых поддерживаемые модели стенограммы для пользователей, основанные на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 3, | |
| ENST00000435633.5 | RPL14-204 | 966 | 52aa | ENSP00000402476.1 | Нонсенс-опосредованный распад | 9184 918Z TS Транскрипт, в котором лучшая поддерживающая мРНК помечена как подозрительная или поддержка поступает из нескольких EST|||||
| ENST00000461368.1 | RPL14-205 | 170 | Нет белка | — | Ген / транскрипт, не содержащий открытой рамки считывания (ORF). Обработанный транскрипт | — | — | — | TSL 2: расшифровка, в которой лучшая поддерживающая мРНК помечена как подозрительная или поддержка исходит от нескольких EST Уровень поддержки транскрипции (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемые модели стенограммы для пользователей, основанные на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 2, | |
| ENST00000479563.5 | RPL14-208 | 794 | Нет белка | — | Альтернативно сплайсированный транскрипт, предположительно содержащий интронную последовательность относительно других, кодирующих тот же ген, транскриптов Сохраненный интрон | — | — | — | TSL 1: транскрипт, в котором все сплайсинговые соединения поддерживаются по крайней мере одной не подозреваемой мРНК. Уровень поддержки стенограммы (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемых моделей стенограммы для пользователей, основанный на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 1, | |
| ENST00000465325.1 | RPL14-207 | 788 | Нет белка | — | Альтернативно сплайсированный транскрипт, предположительно содержащий интронную последовательность относительно других, кодирующих, транскриптов .Сохраненный интрон | — | — | — | TSL 3: стенограмма, в которой единственной поддержкой является один EST Уровень поддержки транскрипции (TSL) — это метод выделения хорошо поддерживаемых и плохо поддерживаемых -поддерживаемые модели стенограммы для пользователей, основанные на типе и качестве выравниваний, используемых для аннотирования стенограммы. TSL: 3, | |
| ENST00000465280.1 | RPL14-206 | 616 | Нет белка | — | Альтернативно сплайсированный транскрипт, предположительно содержащий интронную последовательность относительно других, кодирующих, транскриптов .Сохраненный интрон | — | — | — | TSL 2: транскрипт, в котором лучшая поддерживающая мРНК помечена как подозрительная или поддержка из нескольких EST Уровень поддержки транскрипта (TSL) является методом для выделить для пользователей хорошо поддерживаемые и плохо поддерживаемые модели стенограммы в зависимости от типа и качества выравниваний, используемых для аннотирования стенограммы. |