%d0%b2%d0%b8%d1%80%d1%83%d1%81%20%d0%ba%d0%be%d0%ba%d1%81%d0%b0%d0%ba%d0%b8 — со всех языков на все языки
Все языкиРусскийАнглийскийИспанский────────Айнский языкАканАлбанскийАлтайскийАрабскийАрагонскийАрмянскийАрумынскийАстурийскийАфрикаансБагобоБаскскийБашкирскийБелорусскийБолгарскийБурятскийВаллийскийВарайскийВенгерскийВепсскийВерхнелужицкийВьетнамскийГаитянскийГреческийГрузинскийГуараниГэльскийДатскийДолганскийДревнерусский языкИвритИдишИнгушскийИндонезийскийИнупиакИрландскийИсландскийИтальянскийЙорубаКазахскийКарачаевскийКаталанскийКвеньяКечуаКиргизскийКитайскийКлингонскийКомиКомиКорейскийКриКрымскотатарскийКумыкскийКурдскийКхмерскийЛатинскийЛатышскийЛингалаЛитовскийЛюксембургскийМайяМакедонскийМалайскийМаньчжурскийМаориМарийскийМикенскийМокшанскийМонгольскийНауатльНемецкийНидерландскийНогайскийНорвежскийОрокскийОсетинскийОсманскийПалиПапьяментоПенджабскийПерсидскийПольскийПортугальскийРумынский, МолдавскийСанскритСеверносаамскийСербскийСефардскийСилезскийСловацкийСловенскийСуахилиТагальскийТаджикскийТайскийТатарскийТвиТибетскийТофаларскийТувинскийТурецкийТуркменскийУдмуртскийУзбекскийУйгурскийУкраинскийУрдуУрумскийФарерскийФинскийФранцузскийХиндиХорватскийЦерковнославянский (Старославянский)ЧеркесскийЧерокиЧеченскийЧешскийЧувашскийШайенскогоШведскийШорскийШумерскийЭвенкийскийЭльзасскийЭрзянскийЭсперантоЭстонскийЮпийскийЯкутскийЯпонский
Все языкиРусскийАнглийскийИспанский────────АймараАйнский языкАлбанскийАлтайскийАрабскийАрмянскийАфрикаансБаскскийБашкирскийБелорусскийБолгарскийВенгерскийВепсскийВодскийВьетнамскийГаитянскийГалисийскийГреческийГрузинскийДатскийДревнерусский языкИвритИдишИжорскийИнгушскийИндонезийскийИрландскийИсландскийИтальянскийЙорубаКазахскийКарачаевскийКаталанскийКвеньяКечуаКитайскийКлингонскийКорейскийКрымскотатарскийКумыкскийКурдскийКхмерскийЛатинскийЛатышскийЛингалаЛитовскийЛожбанМайяМакедонскийМалайскийМальтийскийМаориМарийскийМокшанскийМонгольскийНемецкийНидерландскийНорвежскийОсетинскийПалиПапьяментоПенджабскийПерсидскийПольскийПортугальскийПуштуРумынский, МолдавскийСербскийСловацкийСловенскийСуахилиТагальскийТаджикскийТайскийТамильскийТатарскийТурецкийТуркменскийУдмуртскийУзбекскийУйгурскийУкраинскийУрдуУрумскийФарерскийФинскийФранцузскийХиндиХорватскийЦерковнославянский (Старославянский)ЧаморроЧерокиЧеченскийЧешскийЧувашскийШведскийШорскийЭвенкийскийЭльзасскийЭрзянскийЭсперантоЭстонскийЯкутскийЯпонский
Энтеровирусные инфекции — причины, симптомы, диагностика и лечение
Энтеровирусные инфекции — это группа инфекционных заболеваний, развивающихся при поражении человека вирусами рода Enterovirus, характеризующаяся многообразием клинических проявлений.
Общие сведения
Термином «энтеровирусные инфекции» обозначают различные заболевания, вызываемые энтеровирусами Коксаки и ECHO и протекающие с поражение слизистых оболочек, кожи, мышц, нервной системы. Восприимчивость всеобщая, однако массовые вспышки обычно регистрируются в детских коллективах. Большинство детей переносит хотя бы один эпизод энтеровирусной инфекции в возрасте до 5 лет. В разных регионах процентное соотношение здоровых носителей колеблется в пределах 17-46%.
Энтеровирусные инфекции
Причины
Энтеровирусы — РНК-содержащие вирусы, многие из них патогенны для человека: 23 типа вируса Коксаки А, 6 типов – Коксаки В. В эту же группу входит вирус ЕСНО и с 68-го по 71-й типы энтеровирусов. Все возбудители энтеровирусных инфекций отличаются мелкими размерами (от 15 до 35 НМ), устойчивостью к замораживанию и некоторым химическим дезинфицирующим средствам: эфиру, 70%-му спирту, лизолу. В фекалиях могут сохранять жизнеспособность более полугода.
В эту же группу входит вирус ЕСНО и с 68-го по 71-й типы энтеровирусов. Все возбудители энтеровирусных инфекций отличаются мелкими размерами (от 15 до 35 НМ), устойчивостью к замораживанию и некоторым химическим дезинфицирующим средствам: эфиру, 70%-му спирту, лизолу. В фекалиях могут сохранять жизнеспособность более полугода.
Инактивация энтеровирусов происходит при нагревании до 50°С, при высушивании, действии ультрафиолета, дезинфекции раствором формальдегида (0,3%), хлорсодержащими средствами. Резервуаром и источником энтеровирусных инфекций является больной человек или носитель. Заразность отмечается на протяжении недель и месяцев, пик выделения возбудителя приходится на первые дни клинических проявлений. Энтеровирусы передаются с помощью фекально-орального механизма пищевым путем.
Чаще всего заражение происходит при употреблении в пищу загрязненных вирусами овощей. Вирусы попадают на овощи и фрукты при удобрении необеззараженными сточными водами. Некоторые энтеровирусы выделяются с секретом слизистых оболочек дыхательных путей, что может способствовать реализации аэрозольного механизма передачи. Отмечаются случаи заражения контактно-бытовым путем (загрязненные руки, предметы быта, личной гигиены). В случае заражения энтеровирусной инфекцией беременной женщины возможна вертикальная передача возбудителя ребенку. Естественная восприимчивость человека к энтеровирусам высокая, иммунитет обычно типоспецифический, в редких случаях может отмечаться перекрестный.
Некоторые энтеровирусы выделяются с секретом слизистых оболочек дыхательных путей, что может способствовать реализации аэрозольного механизма передачи. Отмечаются случаи заражения контактно-бытовым путем (загрязненные руки, предметы быта, личной гигиены). В случае заражения энтеровирусной инфекцией беременной женщины возможна вертикальная передача возбудителя ребенку. Естественная восприимчивость человека к энтеровирусам высокая, иммунитет обычно типоспецифический, в редких случаях может отмечаться перекрестный.
Классификация
В связи с полиморфизмом клинической симптоматики единая классификация энтеровирусов отсутствует. В нашей стране применяют классифицирование по формам заболевания: типичным и атипичным. К типичным формам течения энтеровирусной инфекции относят герпангину, эпидемическую миалгию, экзантему, асептический серозный менингит.
Кроме того, заболевание может протекать в катаральной, энцефалической, спинальной (полиомиелитоподобной) форме. К атипичным формам также относится энтеровирусный увеит, нефрит, панкреатит, энцефаломиокардит новорожденных, инаппарантную и малую формы, микст-инфекции.
К атипичным формам также относится энтеровирусный увеит, нефрит, панкреатит, энцефаломиокардит новорожденных, инаппарантную и малую формы, микст-инфекции.
Симптомы энтеровирусных инфекций
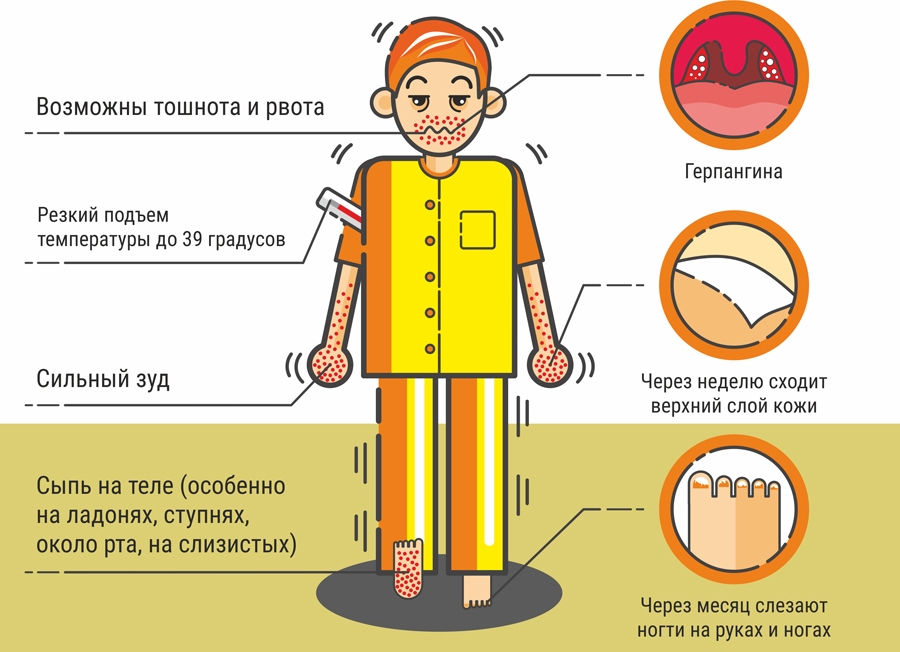
Инкубационный период энтеровирусных инфекций может продолжаться 2-7 дней. Клиника заболевания настолько разнообразна, что можно предположить присутствие нескольких различных патологий. Самые распространенные признаки: интоксикация, повышение температуры, катаральная и абдоминальная симптоматика, полиморфная экзантема.
При остром начале лихорадка может быстро нарастать и достигать 39-40 градусов, после чего сохраняться на протяжении нескольких дней, сопровождается симптомами общей интоксикации (слабость, головная боль, миалгии, возможна рвота и тошнота центрального генеза). Может отмечаться энтеритная диарея. Лихорадка может протекать волнообразно, в особенности при рецидивирующем течении инфекции. При внешнем осмотре отмечают гиперемию лица, инъекции склер, на слизистой ротоглотки отмечают умеренное покраснение, на задней стенке глотки – зернистость. Могут отмечаться разнообразные высыпания.
Могут отмечаться разнообразные высыпания.
Энтеровирусная ангина
Для энтеровирусной герпангины характерна гиперемированность миндалин и небных дужек, возникающая на фоне общей лихорадочной реакции. Вскоре на пораженной слизистой зева образуются герпетоподобные пузырьки, которые лопаются и образуют участки эрозий, покрытые сероватым налетом и имеющие гиперемированный ободок по краю. Пузырьки и эрозии склонны к распространению и слиянию. В течение последующих дней элементы энантемы постепенно заживают. Герпангина нередко сочетается с серозным менингитом.
Эпидемическая миалгия
Эпидемическая миалгия, которую так же называют плевродинией, болезнью Сюльвеста-Финсена и борнхольской болезнью, проявляется, помимо общих признаков инфекции, приступами мышечных болей (являющихся симптомом миозита). Болезненность отмечается в мышцах груди, живота и конечностей. Болевой симптом при энтеровирусном поражении довольно разнообразен по течению. При развитии миозита грудной мускулатуры возможны затруднения дыхания, боль в мышцах брюшной стенки иногда имитирует клинику «острого живота». Как правило, боли длятся около 2-3 дней, после чего стихают. Могут отмечаться случаи рецидива миалгий.
При развитии миозита грудной мускулатуры возможны затруднения дыхания, боль в мышцах брюшной стенки иногда имитирует клинику «острого живота». Как правило, боли длятся около 2-3 дней, после чего стихают. Могут отмечаться случаи рецидива миалгий.
Энтеровирусный менингит
Вирусный менингит представляет собой наиболее распространенную и тяжелую форму энтеровирусной инфекции. Как и всякое воспаление мозговой оболочки, характеризуется положительными менингеальными симптомами: ригидность затылочных мышц, симптомы Брудзинского и Кернига. Пациенты обычно апатичны, вялы, может отмечаться повышенная чувствительность к свету, звукам. Иногда отмечают судороги, психоэмоциональное возбуждение, сознание, как правило, сохранено. Менингеальная симптоматика и повышенная температура тела сохраняется обычно не более 4-7 дней.
Энтеровирусная экзантема
Может как проявляться самостоятельным преимущественным симптомокомплексом, так и сопровождать другие формы энтеровирусных инфекций (серозный менингит, герпангину). Сыпь появляется на 2-3 день лихорадки и интоксикации, локализуется преимущественно на лице, туловище, конечностях, стопах. Сыпь полиморфна, может напоминать экзантему при кори, скарлатине или краснухе, быть розеолезной, пятнисто-папулезной, петехиальной. Часто экзантема сопровождается энантемой в ротовой полости. Могут отмечаться симптомы конъюнктивита, менингизм.
Сыпь появляется на 2-3 день лихорадки и интоксикации, локализуется преимущественно на лице, туловище, конечностях, стопах. Сыпь полиморфна, может напоминать экзантему при кори, скарлатине или краснухе, быть розеолезной, пятнисто-папулезной, петехиальной. Часто экзантема сопровождается энантемой в ротовой полости. Могут отмечаться симптомы конъюнктивита, менингизм.
Энтеровирусная лихорадка
Энтеровирусная инфекция может протекать без каких-либо местных проявлений, единственным симптомом при этом является лихорадка и общая интоксикация. Такую форму еще называют малой болезнью, либо летним гриппом. По типу гриппозного поражения также протекает катаральная (респираторная) форма энтеровирусной инфекции. Лихорадка сопровождается сухим кашлем, саднением в горле и умеренной гиперемией зева, ринитом с серозно-слизистым отделяемым.
Энцефаломиокардит новорожденных
Наиболее тяжело протекает энцефаломиокардит новорожденных. Лихорадка может как присутствовать (обычно имеет двухволновый характер), так и отсутствовать. Отмечается общая выраженная слабость, вялость, сонливость, анорексия, рвота. Кожные покровы цианотичны, одышка, тахикардия. При перкуссии сердца отмечается расширение его границ, при аускультации слышны глухие аритмичные тоны, систолический шум. Печень увеличена, отмечаются отеки. При энцефаломиокардите высока вероятность развития судорожного синдрома, комы. Довольно сложно поддается дифференциальной диагностике с паралитической формой полиомиелита энтеровирусный миелит. Основное отличие – более легкое и кратковременное течение, нередко последующее восстановление двигательной функции.
Отмечается общая выраженная слабость, вялость, сонливость, анорексия, рвота. Кожные покровы цианотичны, одышка, тахикардия. При перкуссии сердца отмечается расширение его границ, при аускультации слышны глухие аритмичные тоны, систолический шум. Печень увеличена, отмечаются отеки. При энцефаломиокардите высока вероятность развития судорожного синдрома, комы. Довольно сложно поддается дифференциальной диагностике с паралитической формой полиомиелита энтеровирусный миелит. Основное отличие – более легкое и кратковременное течение, нередко последующее восстановление двигательной функции.
Энтеровирусная диарея
Вирусный гастроэнтерит характеризуется многократной энтеритной диареей на фоне субфебрилитета, реже – лихорадки. Отмечается боль в животе (преимущественно в правой подвздошной области) со вздутием, может отмечаться рвота. Нередко энтеритная симптоматика сочетается с катаральными явлениями. Может обнаруживаться гепатолиенальный синдром. Заболевание обычно продолжается от нескольких дней до двух недель.
Вирусный конъюнктивит
Для энтеровирусного геморрагического конъюнктивита характерное острое, внезапное начало, отмечается боль в глазах, слезоточивость, светобоязнь. При осмотре конъюнктива гиперемирована, отмечаются многочисленные кровоизлияния, серозные или серозно-гнойные обильные выделения, веки выражено отечны. Первоначально поражается один глаз, позднее заболевание распространяется на второй.
Помимо вышеназванных форм, энтеровирусная инфекция может проявляться в виде энцефалита, безжелтушного гепатита, лимфаденитов различных групп лимфоузлов, поражать сердечную сумку (перикардит), брыжейку кишечника, провоцировать развитие других заболеваний.
Осложнения
Диагностика
Выделение возбудителя из крови, секрета слизистой носоглотки, ликвора или испражнений и вирусологическое исследование возможно, но в широкой клинической практике применяется редко ввиду сложности, длительности и малой диагностической ценности, поскольку выявление и идентификация энтеровируса не всегда является доказательством того, что именно этот возбудитель является этиологической причиной заболевания (в связи с высокой частотой бессимптомного носительства энтеровирусов среди населения).
Основной методикой специфической диагностики энтеровирусов является выявление 4-кратного и большего нарастания титра антител к возбудителю в парных сыворотках. Антитела выявляют с помощью РСК и РТГА.
Лечение энтеровирусных инфекций
Этиотропное лечение для энтеровирусных инфекций не разработано, терапия включает применение дезинтоксикационной терапии и симптоматических средств по показаниям. Тяжелая форма инфекции с поражением нервной системы является показанием к назначению кортикостероидных препаратов и мочегонных средств для коррекции водно-электролитного и кислотно-щелочного баланса. При развитии угрожающих жизни состояний может потребоваться интенсивная терапия и реанимационные меры.
Для стабилизации сосудистого тонуса и улучшения реологических характеристик крови назначают винпоцетин, пентоксифиллин активно применяют витаминотерапию. В зависимости от симптоматики назначают анальгетики, седативные препараты. Развитие вторичной бактериальной инфекции является показанием к назначению курса антибиотикотерапии в соответствии с чувствительностью бактерий к препаратам.
Развитие вторичной бактериальной инфекции является показанием к назначению курса антибиотикотерапии в соответствии с чувствительностью бактерий к препаратам.
Прогноз и профилактика
Энтеровирусные инфекции преимущественно протекают легко, либо в среднетяжелой форме, выздоровление наступает в сроки от нескольких дней, до 2-3 недель. Прогноз ухудшается в случае тяжелого течения, развития осложнений. Некоторые осложнения энтеровирусной инфекции (в основном со стороны нервной системы) могут закончиться летальным исходом, либо оставить после себя неустранимые функциональные нарушения.
Общая профилактика энтеровирусных инфекций включает мероприятия по контролю над загрязненностью объектов окружающей среды канализационными отходами, соблюдением санитарно-гигиенических требований при обезвреживании сточных вод, предоставлением населению пищевых продуктов должной степени эпидемической безопасности.
Индивидуальная профилактика заключается в строгом следовании правилам личной гигиены, необходимой обработке пищевых продуктов. Специфических мер профилактики энтеровирусных инфекций не разработано. Детям до 3 лет, имевшим контакт с больным, назначается интерферон и иммуноглобулин интраназально на 7 дней. В очаге инфекции производится комплекс дезинфицирующих мероприятий.
Специфических мер профилактики энтеровирусных инфекций не разработано. Детям до 3 лет, имевшим контакт с больным, назначается интерферон и иммуноглобулин интраназально на 7 дней. В очаге инфекции производится комплекс дезинфицирующих мероприятий.
В чем опасности вируса Эпштейн-Барр
+ A —
К 40 годам его носителями становятся до 97% людей

Вирус Эпштейн-Барр (ВЭБ) один, но болезней, которые он вызывает, множество. Чаще всего это мононуклеоз. При нем резко повышается температура тела, болит горло, увеличиваются лимфоузлы, печень, селезенка и меняется состав крови. Нередко ВЭБ приводит к ангине. «Часто врачи не знают, с какой ангиной имеют дело — со стрептококковой или вирусной, которую антибиотиками лечить ни в коем случае нельзя», — рассказывает заведующий лабораторией иммунологии и вирусологии ВИЧ-инфекций Александр Семенов.
Еще реже диагностируется причина синдрома хронической усталости — его тоже часто вызывает вирус Эпштейн-Барр. А еще ВЭБ может привести к развитию гепатита и некоторых злокачественных заболеваний.
Открыт этот вирус был совсем недавно, в 1964 году, английским вирусологом Марком Эпштейном в содружестве с его ассистенткой Ивонной Барр. «ВЭБ принадлежит к семейству герпесов, его даже называют герпесом четвертого типа (а всего их 8). Говорят, что XXI век — век герпетических инфекций.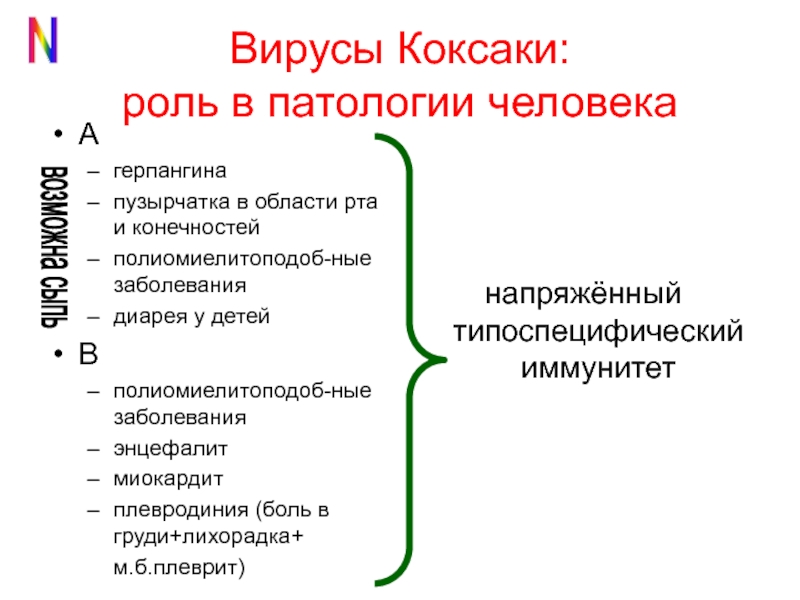
Передается ВЭБ элементарно — с любыми человеческими жидкостями (потом, мокротой, слюной и пр.), а также воздушно-капельным путем. В итоге человек становится вирусоносителем на всю оставшуюся жизнь — вирусы герпеса прячутся в нервных тканях. «Наличие антител при любом герпесе говорит не о защищенности (как, например, при краснухе), а об инфицированности. О том, что вирус внутри нас и мы заразны и любое ослабление иммунитета может обернуться развитием даже хронических заболеваний», — отмечает доктор Семенов.
К подростковому возрасту с ВЭБ успевают познакомиться 50% детей, а к взрослому — почти все. При этом первая встреча в большинстве случаев проходит… без симптомов. Однако стоит подхватить какую-нибудь ОРВИ — и вперед.
Узнать о том, инфицированы вы или нет, сегодня достаточно просто. Специальные анализы крови показывают, есть ли у вас антитела к определенным иммуноглобулинам. Так можно узнать, болен ли человек, может ли он заболеть в ближайшее время, является ли он носителем вируса.
Конечно, всем подряд такая диагностика не нужна. Однако в некоторых случаях она просто необходима. Например, при боли в горле, которая сопровождается увеличением лимфоузлов по всему телу.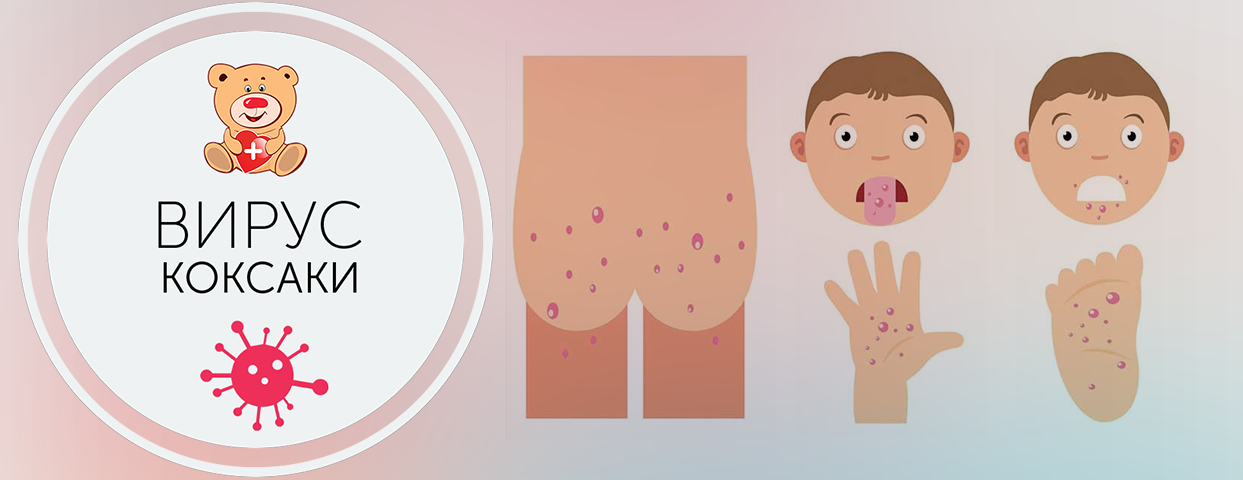 Хотя бы для того, чтобы избежать ненужного приема антибиотиков.
Хотя бы для того, чтобы избежать ненужного приема антибиотиков.
— Ребенку с подозрением на ангину педиатр наверняка выпишет антибиотик. А это вызовет т.н. пенициллиновую реакцию на мононуклеоз — пациент покроется густой малиновой сыпью. В 60-е годы прошлого века это был один из главных диагностических признаков, что это не бактериальная, а вирусная ангина, — рассказывает Александр Семенов.
Между тем заподозрить вызванную ВЭБ ангину достаточно просто — в отличие от бактериальной, при ней лифмоузлы увеличиваются по всему телу, а не только под челюстями, плюс увеличиваются печень и селезенка. «Но часто ли встретишь врача, который при ангине будет щупать печенку?» — вопрошает наш эксперт.
Лечить же такие инфекции сегодня практически нечем. Специальные противовирусные лекарства назначаются только в тяжелых случаях, так как чреваты побочными эффектами. Так что врачи советуют соблюдать постельный режим, пить много жидкости и не переусердствовать с сосудосуживающими препаратами.
Опубликован в газете «Московский комсомолец» №26828 от 6 июня 2015
Заголовок в газете: Вирус замедленного действия
В Коломну пришел вирус Коксаки? – Воскресенск
Заболевание, вызванное вирусом Коксаки, способно привести к смерти. Оно наносит мощнейшие удары по сердцу, печени, поджелудочной железе, поражает все слизистые оболочки организма, включая глаза. Распространяется инфекция просто: по воздуху и через касания. И вот, по мнению родителей дошкольников, распространивших эту информацию в соцсети «ВКонтакте», такое заболевание появилось в Коломне. «Ять» выясняла, стоит ли коломенцам опасаться эпидемииВот что пишет «ВКонтакте» некая Екатерина: «Девочки, привет! А у нас в саду печалька — дети болеют вирусом Коксаки. Хозяин ног находится на Чкалово, и из городских детских садов за день поступили еще шесть детишек» (авторская орфография частично сохранена). Екатерина прикрепила к посту фотографию ног ребенка, лечащегося, как утверждает женщина, в детской больнице на улице Чкалова. На стопах отчетливо видны странные пятна.
Екатерина прикрепила к посту фотографию ног ребенка, лечащегося, как утверждает женщина, в детской больнице на улице Чкалова. На стопах отчетливо видны странные пятна.
Другой пост в Интернете сделала коломчанка Алла. По словам женщины, ее дочь заразилась в детском саду странной инфекцией. В своем обращении к другим родителям Алла подчеркнула, что не хочет наводить панику, однако посоветовала проявить бдительность. «У меня малая ходила в садик, обсалютно здоровая девочка! — пишет женщина. — У ребенка поднялась высокая температура, 39 градусов, сбить не могла и вызвала «скорую». Дочери сделали укол и поставили диагноз — ангина» (авторская орфография частично сохранена).
Коломчанка подробно описывает симптомы заболевания: «Поднимается высокая температура, горло покрывается язвами, высыпание, как во время ветрянки. Эта гадость не лечится антибиотиками! Это вирус! Нужно давать противовирусные препараты. Самое страшное в этом — последствия — менингит. Короче говоря, вирус бушует, не перепутайте его с другими болезнями. Будьте внимательны к детям».
Будьте внимательны к детям».
Как пишет Алла, позже она узнала, что в саду заболели еще несколько детей. Ее возмутило, почему их родители ничего не сказали. «Тупо промолчали!» — констатирует она.
Один случай энтеровирусной инфекции зафиксирован
Чтобы прояснить ситуацию, мы обратились к заместителю начальника территориально отдела Роспотребнадзора по Московской области Владимиру Гурову. «Коксаки относится к энтеровирусной инфекции, — говорит специалист. — Однако в нашей лабораторной службе не определяют ее типы, то есть Коксаки это или нет. В Коломне, конечно, можно сдать анализы на выявление этого вируса, но делают их в Москве, например, в Центральном НИИ эпидемиологии».
В. Гуров сообщил, что в Коломне официально зарегистрирован один случай энтеровирусной инфекции. «Если вы увидели подозрительные пятна на коже ребенка, сразу обращайтесь к врачу, — советует специалист. — Надо ли сдавать анализ, может определить педиатр».
От редакции. Мы направили официальный запрос на электронный адрес КЦРБ с просьбой подтвердить или опровергнуть информацию о случаях заболевания, вызванных вирусом Коксаки, в Коломне. Как только ответ поступит в редакцию, мы его опубликуем.
Как только ответ поступит в редакцию, мы его опубликуем.
Вирусы Коксаки — несколько серотипов РНК-содержащих энтеровирусов, которые хорошо размножаются в желудочно-кишечном тракте. 29 серотипов вирусов Коксаки относят к трем видам энтеровирусов: Enterovirus A, В и С.
Вирусы Коксаки типа A обычно инфицируют кожу и слизистые оболочки, вызывают острый геморрагический конъюнктивит, энтеровирусный стоматит, заболевания верхних дыхательных путей и асептический менингит, миалгию, а также заболевания горла (герпангина). Вирусы Коксаки типа В инфицируют сердце, плевру, печень и поджелудочную железу, вызывают миокардит, перикардит и гепатит. Вирусы Коксаки являются одной из основных причин возникновения асептического менингита.
Как передаются. Чаще всего люди заражаются вирусами Коксаки через загрязненные продукты, предметы быта, воду. Но инфекция передается и воздушно-капельным путем от вирусоносителя здоровому человеку, поэтому ее вспышки бывают не только в бедных, но и во вполне благополучных странах мира. Наиболее подвержены заражению дети в возрасте примерно до 10 лет. Взрослые болеют редко.
Наиболее подвержены заражению дети в возрасте примерно до 10 лет. Взрослые болеют редко.
Симптомы заболевания. У пациента повышается температура до 39-40°С, возможны многократная рвота, диарея, появление сыпи на руках, ногах, груди и лице. Пациент ощущает головную боль, слабость. Поражается слизистая глотки, что затрудняет прием пищи.
По информации ru.wikipedia.org
Вспышки инфекции
В 1997 году эпидемия вируса Коксаки произошла в Малайзии, погибли 30 детей. В апреле 2002 года в связи с распространением вируса (штамм B) были закрыты все школы в Греции, тогда было инфицировано 46 человек, трое скончались. В 2007 году случилась вспышка инфекции в Восточном Китае: погибли 22 ребенка, были заражены более 800 человек, 200 детей госпитализированы.
В 2017 году вирус Коксаки разгулялся на турецких курортах, а под занавес отпускного сезона начал шествие и по России. Тогда Роспотребнадзор открыл электронную «горячую линию» в связи с возможной вспышкой энтеровирусной инфекции на курортах Турции.
что известно, как заразиться, профилактика
В Роспотребнадзор поступило более 500 жалоб на ухудшение здоровья от российских туристов, отдыхавших в Турции. Виной тому вирусы Коксаки, однако, турецкое правительство опровергает эти данные. Главное санитарное ведомство признало небезопасной для россиян обстановку в южных провинциях Турции.
Профессор кафедры инфекционных болезней и эпидемиологии МГМСУ им. А.И. Евдокимова Ирина Шестакова в беседе с корреспондентом Федерального агентства новостей рассказала все, что рядовому гражданину следует знать о вирусах Коксаки, и чем они опасны.
«Если мы говорим о вирусах Коксаки, то они относятся к энтеровирусным инфекциям. Они очень устойчивы к внешней — окружающей среде, хорошо переносят даже низкие температуры. Если мы говорим конкретно о воде, то в свое время Роспотребнадзор опубликовал данные: в водопроводной воде вирус может жить до трех недель; более месяца в обычной речной; сохраняется в соленой воде, во всех бассейнах и так далее — в течение месяца, а то и до двух», — комментирует для ФАН Ирина Шестакова.

«Случаи заболевания отмечались в отелях городов Сиде, Аланья, Белек, Кемер и других. Наибольшее число заболевших — дети до 14 лет», — говорится в сообщении санитарного ведомства.
Как можно заразиться, симптомы вирусов Коксаки
Источником всегда является либо человек, у которого клинически выражено заболевание, либо природная среда.
«Человек может заразиться водным, контактно-бытовым и даже воздушно-капельным путем. Вода, овощи, фрукты, грязные руки, игрушки, столы, вилки, стулья, ручки в туалетах — опасно все, чего мы касаемся руками, а потом тащим в рот, — говорит Шестакова. — Турция полностью не открывает своих данных. То, что было показано по телевизору, это достаточно легкое течение заболевания. Болезнь очень часто проходит под английскими словами в переводе как: «рука, нога, рот». Высыпания появляются на ладонях, стопах, в ротовой полости, зачастую образуется диспепсический синдром (функциональное нарушение кишечного пищеварения, сопровождающееся сосредоточенными в подложечной области болями и ощущением дискомфорта в кишечнике).
На самом деле, поражены могут быть и центральная, и периферическая нервная система, сердце, легкие, печень, почки — все что угодно, даже органы зрения».
 На самом деле, поражены могут быть и центральная, и периферическая нервная система, сердце, легкие, печень, почки — все что угодно, даже органы зрения».
На самом деле, поражены могут быть и центральная, и периферическая нервная система, сердце, легкие, печень, почки — все что угодно, даже органы зрения».Опасность этих вирусов заключается в том, что они могут напоминать банальную кишечную инфекцию. А в определенных случаях, Коксаки может осложниться. Пока от Турции нет данных, говорить, что и как — сложно, считает собеседница ФАН.
«Если говорить по России, то на протяжении последних лет ситуация периодически осложняется. Кажется, в 2011 году была крупная вспышка подъема заболеваемости. Роспотреб подводил итоги 2016 года: по сравнению с 2015-м заболеваемость энтеровирусами подскочила в 1,8 раза. Чаще всего поражает именно детей до 17 лет. Именно на них приходится 90% от всех заболевших. В вирусном ряду Коксаки чаще всего доминирует Коксаки А-6 — то, что вы видели в сюжетах: сыпи — экзантемы, и герпангина в ротовой полости. Есть еще много вирусов, которые вызывают энтеровирусную инфекцию, продолжает рассказ Ирина Шестакова.
 — Я пока не обладаю данными, чем именно вызвана инфекция в Турции: или моноинфекция — только Коксаки А-6, или это вирусы ECHO — ECHO-9, ECHO-30, ECHO-33. В России с января по июнь 2017 года Роспотреб давал 1200 случаев, как наш родной, российский, так и привезенный из той же Турции».
— Я пока не обладаю данными, чем именно вызвана инфекция в Турции: или моноинфекция — только Коксаки А-6, или это вирусы ECHO — ECHO-9, ECHO-30, ECHO-33. В России с января по июнь 2017 года Роспотреб давал 1200 случаев, как наш родной, российский, так и привезенный из той же Турции».Откуда взялись вирусы Коксаки?
Вирусы Коксаки (лат. Coxsackievirus) — несколько серотипов РНК-содержащих энтеровирусов, которые хорошо размножаются в желудочно-кишечном тракте. На сегодняшний день 29 серотипов вирусов Коксаки относят к трем видам энтеровирусов: Enterovirus A, В и С. Эти вирусы стали одной из причин возникновения асептического менингита.
Первоначально эти вирусы были описаны в трудах Гилберта Даллдорфа в 1948-1949 годах. Он и его коллега Грейс Сиклс искали лекарство от полиомиелита. Проводя на мышах опыты, Даллдорф пытался «выявить инфекционный агент из стула больных полиомиелитом, дающий защиту от полиомиелита»
Новое семейство вирусов получило название Коксаки по названию городка в штате Нью-Йорк, где были выделены первые штаммы. Самым страшным случаем, описанным в истории, на сегодняшний день является вспышка вирусов в Восточном Китае, случившаяся в 2007 году. Тогда погибли 22 ребенка, более 800 человек были заражены, 200 детей были госпитализированы в больницы.
Самым страшным случаем, описанным в истории, на сегодняшний день является вспышка вирусов в Восточном Китае, случившаяся в 2007 году. Тогда погибли 22 ребенка, более 800 человек были заражены, 200 детей были госпитализированы в больницы.
«Все зависит от формы вируса. Отдельно идет учет случаев смертности по всем странам, в том числе и по России. Это энтеровирусные менингит и энтеровирусный менингоэнцефалит. Возможно достаточно тяжелое поражение центральной нервной системы. Если заболевание протекает в тяжелой форме, с осложнениями, то понятно, что болезнь может закончиться и летальным исходом. Но это бывает не столь часто. К счастью, болезнь чаще всего протекает в легкой форме, но приятно мало», — констатировала профессор кафедры инфекционных болезней и эпидемиологии МГМСУ им. А.И. Евдокимова Ирина Шестакова.
Профилактика вирусов Коксаки
Специфической профилактики, как утверждает собеседница ФАН, нет.
«Нет, если имеется в виду вакцина.
 Профилактика идет как для всех кишечных инфекций: личная гигиена — не просто вымыть руки, а именно с мылом. Во время вспышки нужно пить только бутилированную воду, налитую в чистейший стакан, лучше всего, в одноразовый. Важно соблюдать температурные условия хранения пищи, тщательно мыть продукты, которые вы употребляете без термической обработки. Нужно оградить детей в большей степени, и взрослых, конечно, от посещения бассейнов до стабилизации ситуации, — подчеркивает Шестакова. — Эта вода меняется не каждый день, а вирус в ней хранится очень долго. Учитывая, что есть еще воздушно-капельный путь, если кто-то на вас чихнет, вы заболеете. Если в Турции предпримут серьезные меры, то заболевание пойдет на спад. Если нет — оно будет разрастаться».
Профилактика идет как для всех кишечных инфекций: личная гигиена — не просто вымыть руки, а именно с мылом. Во время вспышки нужно пить только бутилированную воду, налитую в чистейший стакан, лучше всего, в одноразовый. Важно соблюдать температурные условия хранения пищи, тщательно мыть продукты, которые вы употребляете без термической обработки. Нужно оградить детей в большей степени, и взрослых, конечно, от посещения бассейнов до стабилизации ситуации, — подчеркивает Шестакова. — Эта вода меняется не каждый день, а вирус в ней хранится очень долго. Учитывая, что есть еще воздушно-капельный путь, если кто-то на вас чихнет, вы заболеете. Если в Турции предпримут серьезные меры, то заболевание пойдет на спад. Если нет — оно будет разрастаться».Coxsackievirus — Википедия, бесплатная энциклопедия
El coxsackievirus , или вирус Коксаки, является вирусом, относящимся к семейству Picornaviridae , генералом Enterovirus.
Дебе су номер в городе Коксаки, [1] en el estado de Nueva York (Estados Unidos), donde fue aislado por primera vez.
Разделение на группы:
- Коксаки А.
- Коксаки Б.
Se dividieron sobre la base de ciertas diferencias biológicas y se subdividen en serotipos designados por números sobre la base de diferencias antigénicas adicionales.
Эпидемиология[править]
Los Coxsackievirus реконоценные рецепторы, экспрессирующиеся в различных типах целлюласа и техидоса, вызывающие обширный реперторио де энфермедадес.
Enfermedades[править]
Algunas enfermedades causadas por вирус Коксаки сын:
Предупреждение и контроль[править]
Нет существующих вакунов пара лос вирус Коксаки . Es probable Que la Transmisión se pueda reducir mediante mejoras en la higiene y las condiciones de vida.
Enfermedades, diagnóstico y tratamiento[править]
Enfermedad mano-pie-boca[править]
También llamada fiebre aftosa humana, es el sindrome más conocido provocado por el virus Coxsackie. Общие сведения о серотипе A16. Suelen padecerla mayormente los niños/as pequeños/as de entre 6 a 13 años de edad, aunque también puede aparecer en Adultos, вероятно, нет препятствий для мужчин.
Общие сведения о серотипе A16. Suelen padecerla mayormente los niños/as pequeños/as de entre 6 a 13 años de edad, aunque también puede aparecer en Adultos, вероятно, нет препятствий для мужчин.
La enfermedad no se propaga a partir de animales domésticos o mascotas, pero sí lo puede hacer de una persona a otra.Se puede contraer si se entra en contacto directo con fluos corporales portadores del virus: secreciones de nariz y garganta, слюна, жидкость де ампулы или лас heces де уна персона infectada. El Periodo más contagioso es la primera semana en que se tiene la enfermedad.
Los síntomas привычными, сокращенными а-ля aparicion de pequeñas ampollas en la Palma de las Manos y/o en la planta de los pies, y llagas, igualmente pequeñas en la boca. También puede aparecer fiebre, dolor de cabeza y/o perdida de apetito.
Нет специальных средств, предназначенных для борьбы с инфекцией, которые могут быть вызваны инфекционными заболеваниями.
Evitar alimentos y bebidas calientes о tibios, evitar la sal en lo absoluto.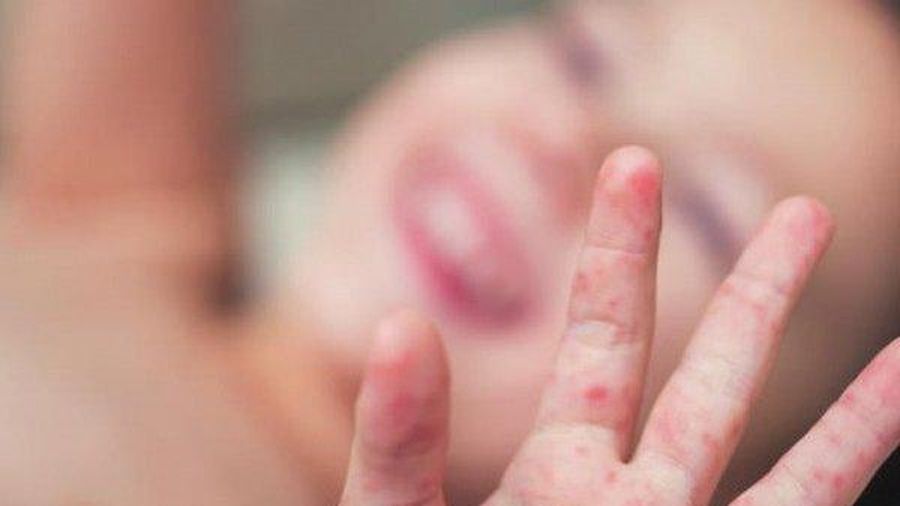 Se debe asegurar Que el enfermo tome suficiente líquido y evitar las comidas picantes, ácidas o que requieran masticar mucho.
Se debe asegurar Que el enfermo tome suficiente líquido y evitar las comidas picantes, ácidas o que requieran masticar mucho.
La enfermedad es muy contagiosa, sobre todo en tempranas edades. Puede contagiarse си уна ampolla се rompe у се entra en contacto кон эль líquido, пор ла слюна у también пор лас heces.
Полное восстановление после 5 и 7 дней.
Оникомадезис[править]
Onicomadesis состоит из ан ла separación o el despegamiento indoloro y sin inflamación de la lamina del lecho ungueal en la zona proximal, apareciendo por debajo una uña nueva. Puede afectar tanto a las uñas de los dedos de las manos como a las de los pies.
Las causas pueden ser variadas: locales (повторяющиеся травмы) и generales, entre las cuales se reconoce la infección por virus Coxsackie.En este caso suele ser autolimitada y de evolución благоприятное. [2] [3]
Справочники[править]
- ↑ «Город Коксаки, администрация, услуги, протоколы, информация сообщества».
- ↑ a b Meseguer Yebra P, Meseguer Yebra C. «Cuando las uñas se caen. La onicomadesis.» Rev Pediatr Aten Primaria . 2013;15:161.e67-e70.
- ↑ Вильяльба Кастаньо, К., Кантареро Вальехо, доктор медицины, Де ла Торре Сесилия, К.. P-12. «Onicomadesis en una consulta de Pediatría de Atención Primaria». Rev Pediatr Aten Primaria . 2010;12(Дополнение 19):e95.
Библиография[править]
- Мюррей П., Розенталь К., Кобаяши Г., Пфаллер М. (1999). Медицинская микробиология . Второе издание, страницы 58, 726, 615, 607, 610, 612, 614.
- «Медлайнплюс, «Энфермедад Мано-пи-бока». Медицинская энциклопедия ».
- «Министерство здравоохранения штата Нью-Йорк.»
- «ProMed Mail: ONICOMADESIS, BROTE ESCOLAR — ESPAÑA (VALENCIA). (загрузить список доступных в Интернет-архиве; добавить историческую, первую и последнюю версии)».
Picornaviridae es una familia de virus infectivos para animales. Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. Генома ARN представляет собой обычное соединение белка с терминалом 5′, которое используется для инициализации транскрипции для полимеризации ARN.Имя, производное от «пико», имеет значение pequeño, напротив «пикорнавируса» буквально означает «вирус ARN pequeños». Представляет собой капсид, содержащий вирусную энволтуру и структурную дефиниду по симетрии икосаэдрики, де ип таманьо де 22 до 30 нм; y por ensamblar los viriones maduros en el citoplasma como compartimento celular. [1]
Contienen un genoma ARN monocatenario positivo y por lo tanto se incluyen en el Grupo IV de la Clasificación de Baltimore. Генома ARN представляет собой обычное соединение белка с терминалом 5′, которое используется для инициализации транскрипции для полимеризации ARN.Имя, производное от «пико», имеет значение pequeño, напротив «пикорнавируса» буквально означает «вирус ARN pequeños». Представляет собой капсид, содержащий вирусную энволтуру и структурную дефиниду по симетрии икосаэдрики, де ип таманьо де 22 до 30 нм; y por ensamblar los viriones maduros en el citoplasma como compartimento celular. [1]
Пикорнавирус, включающий важные патогены для человека и животных. [2] Las enfermedades que causa son variadas, como el resfriado común, полиомиелит и хронические инфекции в Ганадо.Dos categorías maines son los Энтеровирус и Риновирус .
Los Энтеровирус infectan al tracto entérico, mientras que los Риновирусы infectan mainmente nariz y garganta. Los primeros se replican a 37 °C, mientras que los segundos crecen mejor a 33 °C, ya que esta es la Temperature inferior de la nariz. Los Энтеровирус сын estables bajo condiciones ácidas y, por tanto, son capaces de sobrevivir a la exposición al ácido gástrico.Por el contrario, лос Rhinoviruses сына inestables аль ácido у пор эста razón себе limitan nariz у garganta.
Ла-капсайд эс-ун-аррегло-де-60 protómeros en una estructura icosaédrica altamente empaquetada. Cada protómetero consta de 4 polipéptidos denominados VP (proteínas viruses) 1, 2, 3 y 4. Todos estos polipéptidos VP se originan partir de un protómero denominado VP0 que se разделить para dar lugar a los diferentes componentes de la capside. La estructura icosaédrica tiene un número de triangulación 3 puesto que cada uno de los 60 triángulos que componen la cápside se construyen con 3 pequeños triángulos con una subunidad en la esquina.
Dependiendo del typeo y el grado de deshidratación de la partícula virus el diámetro mide alrededor 27-30 нм в диаметре. Вирусная генома имеет долготу на расстоянии 2500 нм, что подтверждается тем, что этот процесс завершается тем, что он совершенствуется в единстве капсида, который содержит сказки, связанные с iones de sodio, и завершает отмену las cargas negativas del ARN causadas por los grupos del fosfato. .
La enfermedades que causanla rubila
Вирусная генома имеет долготу на расстоянии 2500 нм, что подтверждается тем, что этот процесс завершается тем, что он совершенствуется в единстве капсида, который содержит сказки, связанные с iones de sodio, и завершает отмену las cargas negativas del ARN causadas por los grupos del fosfato. .
La enfermedades que causanla rubila
Пикорнавирус содержит единственную филаментную нить ARN de sentido positivo de longitud comprendida entre 7,2 y 9,0 kb de longitud.Como la mayoría de los genomas de ARN de sentido positivo, el material genetico por sí solo es infeccioso, aunque mucho menos virulento que si figura dentro de la partícula вирусная. El genoma es del mismo sentido que el ARNm de los mamíferos, siendo leído desde el extremo 5′ al 3′. Al igual que estos, tiene una cola de poli A en el extremo 3′. Грех эмбарго, разница ARNm де лос mamíferos, лос пикорнавирус не tienen un cap en el extremo 5 ‘, sino una una proteína codificada virusmente denominada VPg.
Hay una región no-traducible (UTR) en ambos extremos del genoma de los picornavirus. El UTR 5 ‘es mayor, en torno al 600-1200 nucleótidos de longitud, en comparación con el UTR 3’, que es de alrededor de 50-100. Se Cree Que el UTR 5 ‘es Importante en la traducción y el 3′ la síntesis de la cadena negativa. Грех эмбарго, el extremo 5′ puede también tener un papel en la virulencia del virus. El resto del genoma codifica proteínas estructurales en el extremo 5’y proteínas no estructurales en el extremo 3’ en una unica poliproteina.
El UTR 5 ‘es mayor, en torno al 600-1200 nucleótidos de longitud, en comparación con el UTR 3’, que es de alrededor de 50-100. Se Cree Que el UTR 5 ‘es Importante en la traducción y el 3′ la síntesis de la cadena negativa. Грех эмбарго, el extremo 5′ puede también tener un papel en la virulencia del virus. El resto del genoma codifica proteínas estructurales en el extremo 5’y proteínas no estructurales en el extremo 3’ en una unica poliproteina.
Experimentos del typeo de curva de crecimiento de un solo paso, разрешающий алос-циентификос вер ла репликацию пикорнавирусов в крупных деталях. La replicación completa se product en el citoplasma de la célula huésped y la infección puede ocurrir incluso en células que no contienen un núcleo (celulas anucleadas) y en las tratadas con actinomicina D (антибиотик ингибирует репликацию вируса, си еста сам производит в эль ядро).
Копия[править]
Вирусные частицы проникают в рецепторы поверхностных клеток.Esto provoca un cambio conformacional en las proteínas de la capside virus y se liberan ácidos miristicos.![]() Estos ácidos forman un poro en la membrana celular a través del cual se inyecta el ARN. Una vez dentro de la célula, el ARN se libera de la cubierta y la cadena positiva se replica a traves de un ARN intermedio de doble cadena que se forma usando RDRP virus (ARN polimerasa dependiente del ARN). La traducción por los ribosomas de la célula huésped no es iniciada por un cap 5′ G, как обычно, sino que se inicia por un IRES (punto de entrada al interior de la ribosoma).
Estos ácidos forman un poro en la membrana celular a través del cual se inyecta el ARN. Una vez dentro de la célula, el ARN se libera de la cubierta y la cadena positiva se replica a traves de un ARN intermedio de doble cadena que se forma usando RDRP virus (ARN polimerasa dependiente del ARN). La traducción por los ribosomas de la célula huésped no es iniciada por un cap 5′ G, как обычно, sino que se inicia por un IRES (punto de entrada al interior de la ribosoma).
Эль цикло де Vida дель вирус эс muy rápido кон todo эль процесс репликации completado en una media де 8 часов. Sin эмбарго, соло 30 minutos después de la infección inicial, la síntesis de proteínas celulares disminuye casi a cero, esto es, se «desconecta». En las próximas 1-2 horas hay una perdida de marginación de cromatina y homogeneidad en el núcleo, antes de que las proteínas viruses comiencen a ser syntetizadas y aparezca una vacuola cerca del núcleo que poco a poco comienza a extenderse cuando la infección де 3 часа. Después де este tiempo, la membrana plasmática celular se vuelve permeable, y a las 4-6 horas las partículas del virus se ensamblan y pueden a veces verse en el citoplasma. En alrededor де 8 horas, ла célula está efectivamente muerta y se lisa para liberar las partículas viruses.
Después де este tiempo, la membrana plasmática celular se vuelve permeable, y a las 4-6 horas las partículas del virus se ensamblan y pueden a veces verse en el citoplasma. En alrededor де 8 horas, ла célula está efectivamente muerta y se lisa para liberar las partículas viruses.
Se han descrito los siguientes géneros:
Algunas especies de la familia se muestran en la siguiente tabla:
Женерос | Especie (* = особый тип) | Серотипос |
| Энтеровирус (EV) | Enterovirus bovino (BEV) | БЭВ-1, БЭВ-2 |
| Энтеровирус человека А | 17 serotipos incluyendo вирус Коксаки A y энтеровирус | |
| Энтеровирус человека В | 56 serotipos, включая энтеровирус, вирус Коксаки B, эховирус y, вирус везикулярной свинины enfermedad | |
| Энтеровирус человека C | 13 серотипов, включая эндоэнтеровирус и вирус Коксаки A1 | |
| Энтеровирус человека D | ЭВ-68, ЭВ-70, ЭВ-94 | |
| Полиовирус (PV) * | PV-1 (цепа Mahoney), PV-2 (цепа Lansing), PV-3 (P3/Leon/37) | |
| Энтеровирус свиньи (PEV) A | ПЭВ-8 | |
| Энтеровирус свиньи B | ПЭВ-9, ПЭВ-10 | |
| Энтеровирус A del simio | СЭВ-А1 | |
| Риновирус | Риновирус человека А * | 74 серотипа |
| Риновирус человека В | 25 серотипов | |
| Гепатовирус | Вирус гепатита А * | Вирус гепатита А человека, вирус гепатита А дель Симио |
| Вирус энцефаломиелита птиц | | |
| Кардиовирус | Вирус энцефаломиокардита * | Вирус Колумбия СК, вирус Мауса Эльберфельда, менговирус |
| Тейловирус | Virus de la encefalomielitis murina de Theiler, вирус человеческого энцефаломиелита де Вилюйск, вирус энцефаломиелита де ла рата | |
| Афтовирус | Virus de la fiebre aftosa [3] * | |
| Вирус конского ринита A (ERAV) | | |
| Пареховирус | Пареховирус человека (HPEV) * | ВПэВ-1, ВПэВ-2, ВПэВ-3 |
| Вирус Люнган | Пареховирус дель ройдор | |
| Эрбовирус | Вирус конского ларинита B (ERBV) * | ЭРБВ-1, ЭРБВ-2 |
| Кобувирус | Вирус Аити * | |
| Кобувирус бычий | | |
| Тешовирус | Teschovirus porcino * | |
| Тремовирус | Вирус энцефаломиелита птиц * | |
Фуэнтес
| ||
 Медицинские предметные рубрики (МЭШ) . Национальная медицинская библиотека. Консультация от 3 сентября 2007 г.
Медицинские предметные рубрики (МЭШ) . Национальная медицинская библиотека. Консультация от 3 сентября 2007 г. Референции[править]
Enlaces externos[править]
Взаимодействие между коксакивирусной инфекцией В3 и α-синуклеином в моделях болезни Паркинсона
Abstract
Болезнь Паркинсона (БП) — одно из наиболее распространенных нейродегенеративных заболеваний. БП патологически характеризуется гибелью дофаминергических нейронов среднего мозга и накоплением внутриклеточных белковых включений, называемых тельцами Леви или нейритами Леви. Основным компонентом телец Леви является α-синуклеин (α-син). Прионоподобное распространение α-сина стало новым механизмом прогрессирования БП. Этот механизм был исследован для выявления факторов, инициирующих патологию Леви, с целью предотвращения дальнейшего прогрессирования БП. Здесь мы демонстрируем, что инфекция вирусом Коксаки B3 (CVB3) может вызывать образование включений, связанных с α-syn, в нейронах, которые могут действовать как триггер для PD. Тельца включения содержали сгруппированные органеллы, в том числе поврежденные митохондрии с фибриллами α-син.Сверхэкспрессия α-Syn ускоряет образование телец включения и индуцирует более концентрические тельца включения. В мозге мышей, инфицированных CVB3, агрегаты α-син наблюдались в теле клеток нейронов среднего мозга. Кроме того, сверхэкспрессия α-syn способствовала репликации CVB3 и связанной с ней цитотоксичности. Трансгенные мыши α-Syn имели низкую выживаемость, повышенную репликацию CVB3 и гибель нейронов, в том числе дофаминергических нейронов черной субстанции.
Основным компонентом телец Леви является α-синуклеин (α-син). Прионоподобное распространение α-сина стало новым механизмом прогрессирования БП. Этот механизм был исследован для выявления факторов, инициирующих патологию Леви, с целью предотвращения дальнейшего прогрессирования БП. Здесь мы демонстрируем, что инфекция вирусом Коксаки B3 (CVB3) может вызывать образование включений, связанных с α-syn, в нейронах, которые могут действовать как триггер для PD. Тельца включения содержали сгруппированные органеллы, в том числе поврежденные митохондрии с фибриллами α-син.Сверхэкспрессия α-Syn ускоряет образование телец включения и индуцирует более концентрические тельца включения. В мозге мышей, инфицированных CVB3, агрегаты α-син наблюдались в теле клеток нейронов среднего мозга. Кроме того, сверхэкспрессия α-syn способствовала репликации CVB3 и связанной с ней цитотоксичности. Трансгенные мыши α-Syn имели низкую выживаемость, повышенную репликацию CVB3 и гибель нейронов, в том числе дофаминергических нейронов черной субстанции. Эти результаты могут быть связаны с различными путями, связанными с аутофагией, задействованными CVB3 и α-syn.Это исследование разъяснило механизм образования телец Леви и патогенез БП, связанной с инфекцией CVB3.
Эти результаты могут быть связаны с различными путями, связанными с аутофагией, задействованными CVB3 и α-syn.Это исследование разъяснило механизм образования телец Леви и патогенез БП, связанной с инфекцией CVB3.
Резюме автора
Прионоподобное распространение α-сина стало новым механизмом, участвующим в прогрессировании болезни Паркинсона (БП). Этот процесс был тщательно исследован для выявления факторов, которые инициируют патологию Леви, чтобы предотвратить дальнейшее прогрессирование БП. Тем не менее начальные триггеры образования телец Леви (ТЛ), приводящие к ускорению процесса, до сих пор остаются неуловимыми.Инфекция все чаще признается фактором риска БП. В частности, сообщалось, что несколько вирусов связаны как с острым, так и с хроническим паркинсонизмом. Было высказано предположение, что периферические инфекции, включая вирусные инфекции, сопровождающие воспаление, могут вызывать БП. В настоящем исследовании мы исследовали, взаимодействует ли вирус коксаки B3 (CVB3) с α-син, чтобы вызвать агрегацию и дальнейшее образование телец Леви, тем самым действуя как триггер, и влияет ли α-син на репликацию вируса Коксаки. Важно определить факторы, которые инициируют патологию Леви, чтобы понять патогенез БП. Наши результаты проясняют механизм образования LB и патогенез БП, связанной с инфекцией CVB3.
Важно определить факторы, которые инициируют патологию Леви, чтобы понять патогенез БП. Наши результаты проясняют механизм образования LB и патогенез БП, связанной с инфекцией CVB3.
Образец цитирования: Park SJ, Jin U, Park SM (2021) Взаимодействие между коксакивирусной инфекцией B3 и α-синуклеином в моделях болезни Паркинсона. ПЛОС Патог 17(10): е1010018. https://doi.org/10.1371/journal.ppat.1010018
Редактор: Джордж А.Белов, Мэрилендский университет, США
Поступила в редакцию: 16 февраля 2021 г.; Принято: 8 октября 2021 г .; Опубликовано: 25 октября 2021 г.
Copyright: © 2021 Park et al. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные содержатся в рукописи и файлах вспомогательной информации.
Финансирование: Это исследование было поддержано грантами Национального исследовательского фонда Кореи (NRF), финансируемыми правительством Кореи (Министерством науки и ИКТ и Министерством образования) (грант № NRF-2017R1E1A1A01073713 SMP, NRF-2019R1A5A2026045 к СМП и М-2021А040300173 к УЖ). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Болезнь Паркинсона (БП) — одно из наиболее распространенных нейродегенеративных заболеваний. БП патологически характеризуется гибелью дофаминергических нейронов среднего мозга и накоплением внутриклеточных белковых включений, называемых тельцами Леви (ТЛ) или нейритами Леви (НЛ) [1, 2]. Основным компонентом этих включений является α-синуклеин (α-син) [3]. Белковые включения с агрегатами α-син также наблюдались при других нейродегенеративных заболеваниях, таких как множественная системная атрофия и деменция с тельцами Леви, которые в совокупности называются α-синуклеинопатиями [4].Мультипликации и миссенс-мутации гена α-syn были выявлены у пациентов с ранним началом семейной БП [5]. Кроме того, полногеномные ассоциативные исследования продемонстрировали сильную связь между геном α-син и спорадической болезнью Паркинсона [6, 7], что свидетельствует о важной роли α-сина в патогенезе БП.
Основным компонентом этих включений является α-синуклеин (α-син) [3]. Белковые включения с агрегатами α-син также наблюдались при других нейродегенеративных заболеваниях, таких как множественная системная атрофия и деменция с тельцами Леви, которые в совокупности называются α-синуклеинопатиями [4].Мультипликации и миссенс-мутации гена α-syn были выявлены у пациентов с ранним началом семейной БП [5]. Кроме того, полногеномные ассоциативные исследования продемонстрировали сильную связь между геном α-син и спорадической болезнью Паркинсона [6, 7], что свидетельствует о важной роли α-сина в патогенезе БП.
Патология Леви сначала проявляется в обонятельных луковицах и дорсальных двигательных ядрах блуждающего нерва, которые связаны с энтеральной нервной системой. Патология постепенно вовлекает все больше областей нервной системы, а затем и области коры по мере прогрессирования заболевания [8].Эта патология, по-видимому, возникает до появления двигательных симптомов при БП и может быть связана с желудочно-кишечными и обонятельными дисфункциями, которые часто наблюдаются в продромальной фазе БП [9]. Существенные экспериментальные данные in vitro и in vivo указывают на то, что прионоподобное распространение α-сина является новым механизмом прогрессирования БП [10–12]. Ориентация на этот механизм может позволить разработать модифицирующую болезнь терапию для пациентов с БП.Однако первоначальные триггеры формирования СП, приводящие к ускорению процесса, остаются неуловимыми.
Существенные экспериментальные данные in vitro и in vivo указывают на то, что прионоподобное распространение α-сина является новым механизмом прогрессирования БП [10–12]. Ориентация на этот механизм может позволить разработать модифицирующую болезнь терапию для пациентов с БП.Однако первоначальные триггеры формирования СП, приводящие к ускорению процесса, остаются неуловимыми.
Вирусная инфекция все чаще признается фактором риска БП. Ряд вирусов ассоциирован как с острым, так и с хроническим паркинсонизмом. Эти вирусы включают вирус гриппа, вирус Коксаки, вирус японского энцефалита В, вирус западного энцефалита лошадей и вирус герпеса [13]. Было высказано предположение, что периферические инфекции, в том числе вирусные инфекции, сопровождающие воспаление, могут вызывать БП [14].
Вирус Коксаки представляет собой одноцепочечный РНК-содержащий вирус, принадлежащий к семейству вирусов Picornaviridae рода Enterovirus [15]. Более 90% инфекций, вызванных вирусом Коксаки, протекают бессимптомно.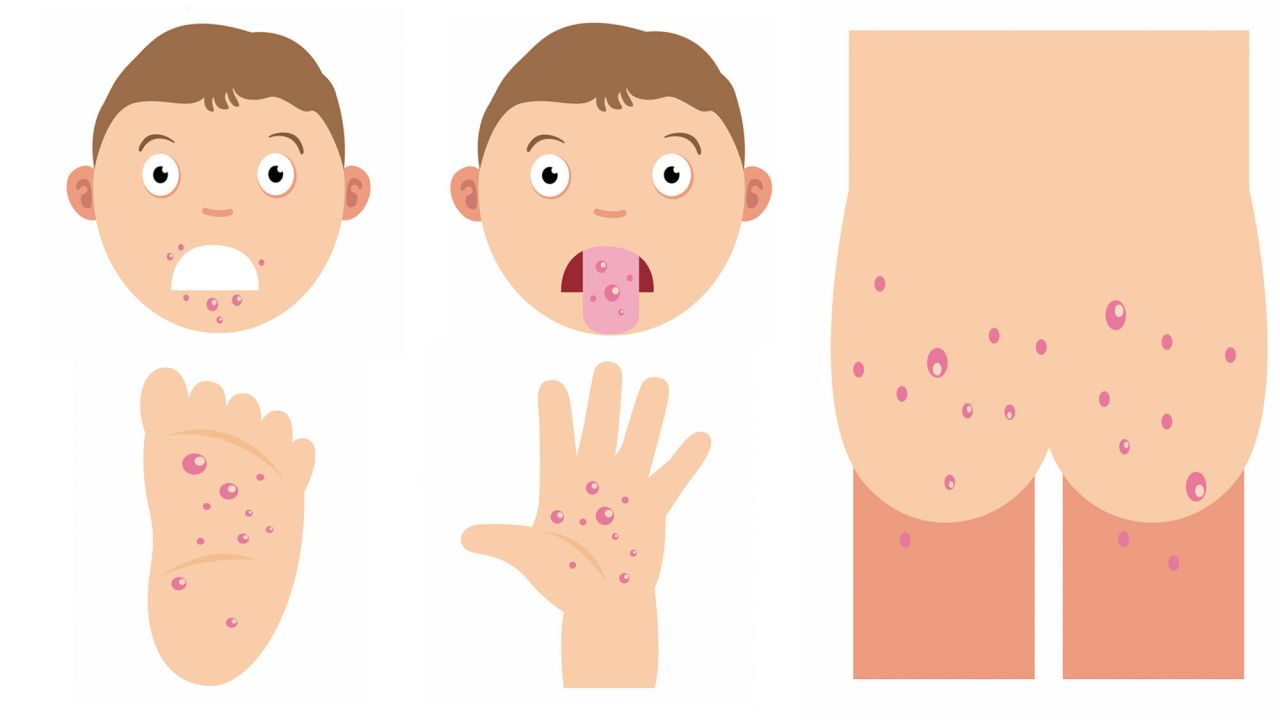 Клинически младенцы или молодые люди легко заражаются этим вирусом, и у некоторых развивается тяжелый миокардит [16] или менингит [17]. Персистирующая инфекция вируса Коксаки также связана с хроническим миокардитом, дилатационной кардиомиопатией [18] и диабетом I типа [19].
Клинически младенцы или молодые люди легко заражаются этим вирусом, и у некоторых развивается тяжелый миокардит [16] или менингит [17]. Персистирующая инфекция вируса Коксаки также связана с хроническим миокардитом, дилатационной кардиомиопатией [18] и диабетом I типа [19].
В недавнем отчете описаны вирусоподобные частицы и антиген энтеровируса в нейронах ствола мозга при БП [20]. Это открытие подтолкнуло к предположению, что энтеровирусная инфекция при БП может служить источником для агрегации α-син в дополнение к прямому цитопатическому эффекту вирусной инфекции на нейроны. Кроме того, α-син ингибирует инфекцию вирусом Западного Нила (ВЗН), действуя как фактор рестрикции вируса [21], что позволяет предположить, что экспрессия α-син может влиять на вирусную инфекцию в центральной нервной системе (ЦНС).
В настоящем исследовании мы исследовали, взаимодействует ли вирус Коксаки B3 (CVB3) с α-syn, вызывая агрегацию и дальнейшее образование LB, и влияет ли α-syn на репликацию вируса Коксаки.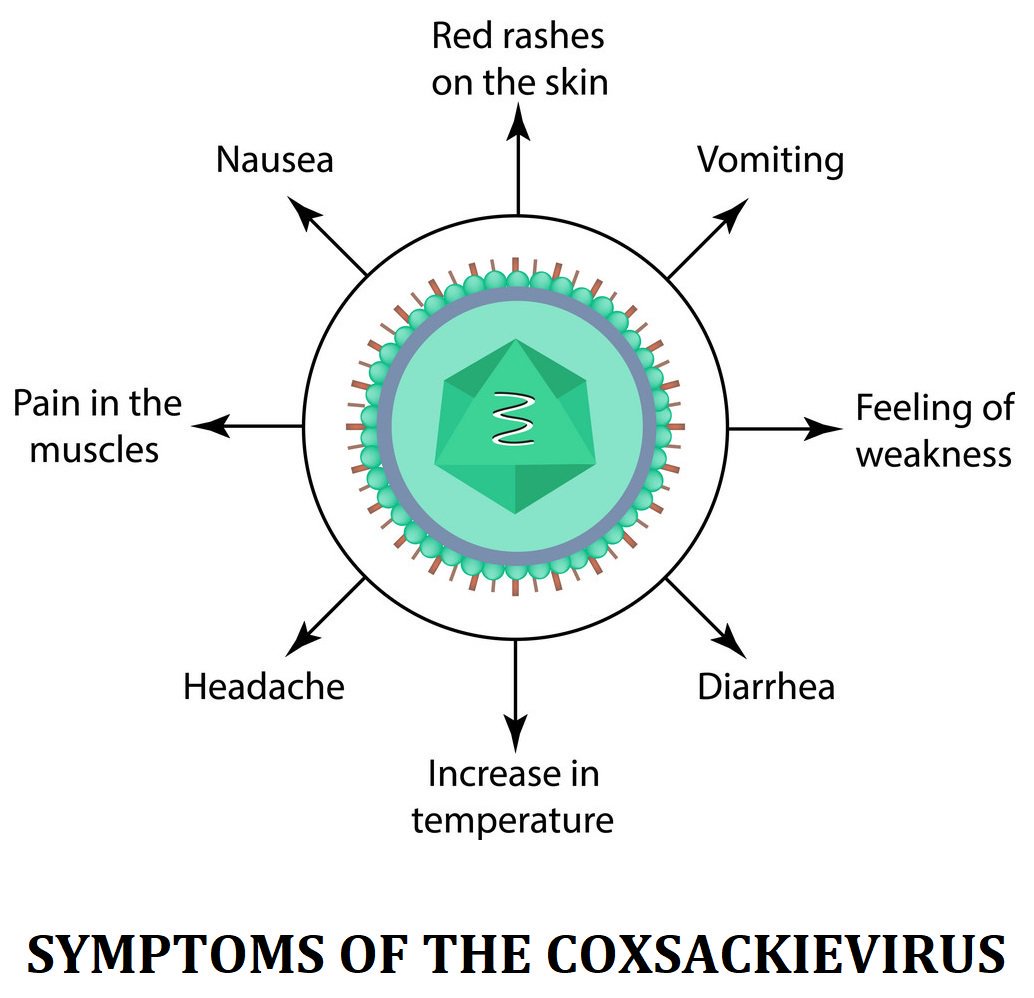
Результаты
Инфекция CVB3 регулирует экспрессию α-син в нейронах
Чтобы выяснить, влияет ли CVB3 на α-syn, мы инфицировали дифференцированные клетки SH-SY5Y (клетки dSH-SY5Y) CVB3 (MOI 0,25) в течение 24 часов. CVB3 VP1 колокализовался с α-сином, и интенсивность α-сина в инфицированных клетках увеличивалась.Интересно, что мы наблюдали, что инфекция CVB3 индуцировала образование больших агрегатов α-син, которые полностью заполняли цитоплазму и оттесняли ядро, создавая вид полумесяца. Этот вид был более выражен в клетках dSH-SY5Y, сверхэкспрессирующих α-syn (рис. 1А). Это не было связано с перекрестной реактивностью α-син антитела с CVB3 (S1A фиг.), и эти включения были эозинофильными (S1B фиг.). В первичных корковых нейронах наблюдалась сходная колокализация VP1 с α-syn (рис. 1В).Напротив, уровни мРНК α-syn были снижены в клетках dSH-SY5Y и первичных нейронах коры, инфицированных CVB3 (рис. 1C). При инфицировании CVB3 не наблюдалось значительной цитотоксичности (рис. S1C), что позволяет предположить, что снижение уровня мРНК α-син может быть не связано с цитотоксичностью. Инфицирование клеток многими вирусами приводит к ингибированию транскрипции или трансляции мРНК клетки-хозяина, что называется выключением хозяина [22, 23]. Уровни нескольких мРНК, включая β-актин, гистон h4 и polr2, которые, как известно, связаны с феноменом отключения [24–26], не изменялись при заражении CVB3.Более того, уровни генов, связанных с БП, таких как DJ-1, PINK1 и паркин, не изменялись при заражении CVB3 (рис. S1D), что позволяет предположить, что снижение уровня мРНК α-син может также не быть связано с отключением хозяина. явление. Анализ базы данных с открытым исходным кодом (GSE 19496) также показал, что уровни мРНК α-syn в CVB3-инфицированном сердце мыши были снижены по сравнению с таковыми в контроле (S1E Fig). Вестерн-блоттинг показал, что эндогенная экспрессия α-син была снижена. Интересно, что экспрессия эктопически сверхэкспрессированного α-syn также была снижена.
S1C), что позволяет предположить, что снижение уровня мРНК α-син может быть не связано с цитотоксичностью. Инфицирование клеток многими вирусами приводит к ингибированию транскрипции или трансляции мРНК клетки-хозяина, что называется выключением хозяина [22, 23]. Уровни нескольких мРНК, включая β-актин, гистон h4 и polr2, которые, как известно, связаны с феноменом отключения [24–26], не изменялись при заражении CVB3.Более того, уровни генов, связанных с БП, таких как DJ-1, PINK1 и паркин, не изменялись при заражении CVB3 (рис. S1D), что позволяет предположить, что снижение уровня мРНК α-син может также не быть связано с отключением хозяина. явление. Анализ базы данных с открытым исходным кодом (GSE 19496) также показал, что уровни мРНК α-syn в CVB3-инфицированном сердце мыши были снижены по сравнению с таковыми в контроле (S1E Fig). Вестерн-блоттинг показал, что эндогенная экспрессия α-син была снижена. Интересно, что экспрессия эктопически сверхэкспрессированного α-syn также была снижена. Это явление, вероятно, было более серьезным с увеличением титра вируса (рис. 1D). Кроме того, когда мы внутрибрюшинно инфицировали мышей WT CVB3, в головном мозге наблюдалось снижение уровней мРНК и белка α-syn (рис. 1E и 1F). Учитывая, что CVB3 не инфицировал все клетки, эти результаты привели нас к предположению, что α-syn может регулироваться по-разному в CVB3-инфицированных клетках и соседних клетках. Чтобы изучить это, мы сравнили уровни α-син в неинфицированных и инфицированных условиях. Повышенные уровни α-син наблюдались в клетках, инфицированных CVB3, и уровни были заметно снижены в клетках рядом с клетками, инфицированными CVB3 (рис. 1G).Аналогичные результаты наблюдались в первичных нейронах коры (рис. 1H), что позволяет предположить, что α-syn по-разному регулировался в CVB3-инфицированных клетках и соседних клетках. При обработке клеток поли IC, искусственным аналогом имитирующей РНК вирусной инфекции [27], экспрессия α-син увеличивалась как на уровне мРНК, так и на уровне белка, тогда как агрегаты α-син не наблюдались (рис.
Это явление, вероятно, было более серьезным с увеличением титра вируса (рис. 1D). Кроме того, когда мы внутрибрюшинно инфицировали мышей WT CVB3, в головном мозге наблюдалось снижение уровней мРНК и белка α-syn (рис. 1E и 1F). Учитывая, что CVB3 не инфицировал все клетки, эти результаты привели нас к предположению, что α-syn может регулироваться по-разному в CVB3-инфицированных клетках и соседних клетках. Чтобы изучить это, мы сравнили уровни α-син в неинфицированных и инфицированных условиях. Повышенные уровни α-син наблюдались в клетках, инфицированных CVB3, и уровни были заметно снижены в клетках рядом с клетками, инфицированными CVB3 (рис. 1G).Аналогичные результаты наблюдались в первичных нейронах коры (рис. 1H), что позволяет предположить, что α-syn по-разному регулировался в CVB3-инфицированных клетках и соседних клетках. При обработке клеток поли IC, искусственным аналогом имитирующей РНК вирусной инфекции [27], экспрессия α-син увеличивалась как на уровне мРНК, так и на уровне белка, тогда как агрегаты α-син не наблюдались (рис. 1I–1K). ). Эти данные свидетельствуют о том, что наблюдения были специфичны для инфекции CVB3. Анализ других данных из открытых источников (GSE 7621) также выявил снижение уровня мРНК α-син в мозге пациентов с БП по сравнению с таковым у здоровых людей (рис. S1F).Эти результаты свидетельствуют о том, что инфекция CVB3 индуцирует большие цитозольные агрегаты, которые локализованы совместно с α-syn, и что экспрессия α-syn по-разному регулируется в инфицированных клетках и соседних клетках.
1I–1K). ). Эти данные свидетельствуют о том, что наблюдения были специфичны для инфекции CVB3. Анализ других данных из открытых источников (GSE 7621) также выявил снижение уровня мРНК α-син в мозге пациентов с БП по сравнению с таковым у здоровых людей (рис. S1F).Эти результаты свидетельствуют о том, что инфекция CVB3 индуцирует большие цитозольные агрегаты, которые локализованы совместно с α-syn, и что экспрессия α-syn по-разному регулируется в инфицированных клетках и соседних клетках.
Рис. 1. Инфекция CVB3 регулирует экспрессию α-syn в нейронах.
(A) Иммуноцитохимические (ICC) изображения клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 (MOI 0,25) (красный) в течение 24 часов. Анализировали интенсивность α-син (зеленый). Значения получены из четырех независимых экспериментов (n = 4).*** P <0,001, однофакторный дисперсионный анализ с тестом множественных сравнений Тьюки. (B) ICC-изображения первичных кортикальных нейронов мыши, инфицированных CVB3 (MOI 5) в течение 24 часов. Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (C) Относительные уровни экспрессии мРНК α-syn между контрольными клетками dSH-SY5Y, инфицированными CVB3 (MOI 0,25), и первичными кортикальными нейронами мыши (MOI 5), инфицированными CVB3, в течение 24 часов. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, непарный t-критерий.(D) Клетки WT и α-syn OE dSH-SY5Y были инфицированы указанными MOI CVB3 в течение 24 часов. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (E) Относительные уровни экспрессии мРНК α-syn между контрольными (n = 6) и CVB3-инфицированными мышами в полушариях головного мозга (n = 14) после 4 дней внутрибрюшинной (IP) инъекции с 1.0 × 10 6 бляшкообразующих единиц (БОЕ) CVB3 в 100 мкл PBS.
Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (C) Относительные уровни экспрессии мРНК α-syn между контрольными клетками dSH-SY5Y, инфицированными CVB3 (MOI 0,25), и первичными кортикальными нейронами мыши (MOI 5), инфицированными CVB3, в течение 24 часов. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, непарный t-критерий.(D) Клетки WT и α-syn OE dSH-SY5Y были инфицированы указанными MOI CVB3 в течение 24 часов. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (E) Относительные уровни экспрессии мРНК α-syn между контрольными (n = 6) и CVB3-инфицированными мышами в полушариях головного мозга (n = 14) после 4 дней внутрибрюшинной (IP) инъекции с 1.0 × 10 6 бляшкообразующих единиц (БОЕ) CVB3 в 100 мкл PBS. ** P <0,01, непарный t-критерий. (F) Относительные уровни экспрессии α-syn между контрольными (n = 3) и инфицированными CVB3 полушариями головного мозга мышей (n = 3) после 7-дневной внутрибрюшинной инъекции 1,0 × 10 6 БОЕ CVB3. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. ** P <0,01, непарный t-критерий. (Г и Н). ICC-изображения контрольных и инфицированных CVB3 клеток WT, α-syn OE dSH-SY5Y (G) и первичных кортикальных нейронов мыши (H), которые были инфицированы CVB3 в течение 24 часов.Анализировали интенсивность α-Syn (зеленый) указанного состояния. Значения получены из трех (G) или четырех (H) независимых экспериментов (n = 3 или 4). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. * P <0,05, *** P <0,001, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (I) Относительные уровни экспрессии α-syn мРНК между контролем и клетками dSH-SY5Y, трансфицированными 1 мкг/мл поли IC, в течение 4 часов.
** P <0,01, непарный t-критерий. (F) Относительные уровни экспрессии α-syn между контрольными (n = 3) и инфицированными CVB3 полушариями головного мозга мышей (n = 3) после 7-дневной внутрибрюшинной инъекции 1,0 × 10 6 БОЕ CVB3. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. ** P <0,01, непарный t-критерий. (Г и Н). ICC-изображения контрольных и инфицированных CVB3 клеток WT, α-syn OE dSH-SY5Y (G) и первичных кортикальных нейронов мыши (H), которые были инфицированы CVB3 в течение 24 часов.Анализировали интенсивность α-Syn (зеленый) указанного состояния. Значения получены из трех (G) или четырех (H) независимых экспериментов (n = 3 или 4). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. * P <0,05, *** P <0,001, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (I) Относительные уровни экспрессии α-syn мРНК между контролем и клетками dSH-SY5Y, трансфицированными 1 мкг/мл поли IC, в течение 4 часов. Значения получены из трех независимых экспериментов (n = 3). ** P <0,01, непарный t-критерий.(J) Клетки WT и α-syn OE dSH-SY5Y, трансфицированные 1 мкг/мл поли IC в течение 4 часов. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). * P <0,05, непарный t-критерий. (K) ICC-изображение контрольных и 1 мкг/мл поли IC-трансфицированных клеток dSH-SY5Y в течение 24 часов. Анализировали интенсивность α-син (зеленый). Значения получены из трех независимых экспериментов (n = 3). Масштабная линейка указывает 20 мкм.Синий указывает на Hoechst. *** P <0,001, непарный t-критерий.
Значения получены из трех независимых экспериментов (n = 3). ** P <0,01, непарный t-критерий.(J) Клетки WT и α-syn OE dSH-SY5Y, трансфицированные 1 мкг/мл поли IC в течение 4 часов. Вестерн-блоттинг проводили с указанными антителами, и уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). * P <0,05, непарный t-критерий. (K) ICC-изображение контрольных и 1 мкг/мл поли IC-трансфицированных клеток dSH-SY5Y в течение 24 часов. Анализировали интенсивность α-син (зеленый). Значения получены из трех независимых экспериментов (n = 3). Масштабная линейка указывает 20 мкм.Синий указывает на Hoechst. *** P <0,001, непарный t-критерий.
https://doi.org/10.1371/journal.ppat.1010018.g001
Инфекция CVB3 индуцирует образование LB-подобных телец включения в нейронах
CVB3 образует очень большие связанные с аутофагией структуры, называемые мегафагосомами, в ацинарных клетках поджелудочной железы мышей, структура которых представляет собой комплексы репликации вируса [28]. Чтобы более подробно исследовать, локализуются ли эти большие агрегаты вместе с α-syn, мы окрашивали клетки на ассоциированный с микротрубочками белок 1A/1B легкой цепи 3B (LC3), маркер аутофагосом [29].Эти структуры полностью колокализованы с LC3 в клетках dSH-SY5Y, сверхэкспрессирующих α-syn (OE) клетках dSH-SY5Y и первичных нейронах коры (рис. 2А). Эти структуры также совместно локализованы с pSer129 α-syn в клетках α-syn OE dSH-SY5Y и первичных нейронах коры (рис. 2B). Колокализация этих агрегатов с убиквитином, другим маркером LB [30], более четко наблюдалась в клетках α-syn OE dSH-SY5Y, что позволяет предположить, что эти структуры могут быть LB-подобными включениями (рис. 2C). При заражении энтеровирусом 71 (EV71), другим вирусом семейства Picornaviridae , клетки dSH-SY5Y образовывали более мелкие LC3-позитивные агрегаты.Однако они не локализовались совместно с α-syn (рис. S2), что позволяет предположить, что образование LB-подобных телец включения, содержащих α-syn, было специфичным для CVB3.
Чтобы более подробно исследовать, локализуются ли эти большие агрегаты вместе с α-syn, мы окрашивали клетки на ассоциированный с микротрубочками белок 1A/1B легкой цепи 3B (LC3), маркер аутофагосом [29].Эти структуры полностью колокализованы с LC3 в клетках dSH-SY5Y, сверхэкспрессирующих α-syn (OE) клетках dSH-SY5Y и первичных нейронах коры (рис. 2А). Эти структуры также совместно локализованы с pSer129 α-syn в клетках α-syn OE dSH-SY5Y и первичных нейронах коры (рис. 2B). Колокализация этих агрегатов с убиквитином, другим маркером LB [30], более четко наблюдалась в клетках α-syn OE dSH-SY5Y, что позволяет предположить, что эти структуры могут быть LB-подобными включениями (рис. 2C). При заражении энтеровирусом 71 (EV71), другим вирусом семейства Picornaviridae , клетки dSH-SY5Y образовывали более мелкие LC3-позитивные агрегаты.Однако они не локализовались совместно с α-syn (рис. S2), что позволяет предположить, что образование LB-подобных телец включения, содержащих α-syn, было специфичным для CVB3. Затем мы исследовали эти структуры с помощью просвечивающей электронной микроскопии (ПЭМ). В отсутствие инфекции CVB3 внутриклеточные органеллы были рассредоточены по всей цитоплазме в клетках dSH-SY5Y и клетках α-syn OE dSH-SY5Y, тогда как органеллы инфицированных вирусом клеток накапливались в сферических структурах (рис. 2D). Эти сферические структуры содержали различные дезорганизованные органеллы, состояли из большого количества везикул, поврежденных митохондрий и аутофагических компонентов (рис. 2D), подобно ранее наблюдаемым мегафагосомам [28].Эти структуры также были аналогичны наблюдавшимся ранее СП [31]. Кроме того, наблюдались кристаллические массивы в форме сот в качестве репликационных частиц CVB3 [32], которые были более распространены в клетках α-syn OE dSH-SY5Y, чем в клетках dSH-SY5Y (рис. 2E). Они также наблюдались в первичных нейронах мыши (рис. 2F). В клетках, инфицированных CVB3, наблюдали фибриллярные структуры. Ширина и длина этих фибриллярных структур в клетках dSH-SY5Y составляли примерно 20 нм и 400 нм соответственно (рис.
Затем мы исследовали эти структуры с помощью просвечивающей электронной микроскопии (ПЭМ). В отсутствие инфекции CVB3 внутриклеточные органеллы были рассредоточены по всей цитоплазме в клетках dSH-SY5Y и клетках α-syn OE dSH-SY5Y, тогда как органеллы инфицированных вирусом клеток накапливались в сферических структурах (рис. 2D). Эти сферические структуры содержали различные дезорганизованные органеллы, состояли из большого количества везикул, поврежденных митохондрий и аутофагических компонентов (рис. 2D), подобно ранее наблюдаемым мегафагосомам [28].Эти структуры также были аналогичны наблюдавшимся ранее СП [31]. Кроме того, наблюдались кристаллические массивы в форме сот в качестве репликационных частиц CVB3 [32], которые были более распространены в клетках α-syn OE dSH-SY5Y, чем в клетках dSH-SY5Y (рис. 2E). Они также наблюдались в первичных нейронах мыши (рис. 2F). В клетках, инфицированных CVB3, наблюдали фибриллярные структуры. Ширина и длина этих фибриллярных структур в клетках dSH-SY5Y составляли примерно 20 нм и 400 нм соответственно (рис. 2G).Фибриллы были более многочисленными и длинными в клетках α-syn OE dSH-SY5Y, чем в клетках dSH-SY5Y, и не были обнаружены в клетках α-syn KO dSH-SY5Y (рис. 2G). Эти паттерны также наблюдались в первичных корковых нейронах мышей WT и α-syn трансгенных (TG) мышей (рис. 2H). Эти результаты позволяют предположить, что инфекция CVB3 индуцирует образование LB-подобных включений в нейронах. Кроме того, поврежденные митохондрии анализировали, как описано ранее [33]. В состоянии покоя не было различий в митохондриальной морфологии между клетками dSH-SY5Y и клетками α-syn OE dSH-SY5Y.Однако после заражения CVB3 количество поврежденных митохондрий увеличивалось в клетках dSH-SY5Y и было еще больше в клетках α-syn OE dSH-SY5Y (рис. 2I и 2J), что позволяет предположить, что повреждение митохондрий при заражении CVB3 ускоряется в ответ на к гиперэкспрессии α-syn.
2G).Фибриллы были более многочисленными и длинными в клетках α-syn OE dSH-SY5Y, чем в клетках dSH-SY5Y, и не были обнаружены в клетках α-syn KO dSH-SY5Y (рис. 2G). Эти паттерны также наблюдались в первичных корковых нейронах мышей WT и α-syn трансгенных (TG) мышей (рис. 2H). Эти результаты позволяют предположить, что инфекция CVB3 индуцирует образование LB-подобных включений в нейронах. Кроме того, поврежденные митохондрии анализировали, как описано ранее [33]. В состоянии покоя не было различий в митохондриальной морфологии между клетками dSH-SY5Y и клетками α-syn OE dSH-SY5Y.Однако после заражения CVB3 количество поврежденных митохондрий увеличивалось в клетках dSH-SY5Y и было еще больше в клетках α-syn OE dSH-SY5Y (рис. 2I и 2J), что позволяет предположить, что повреждение митохондрий при заражении CVB3 ускоряется в ответ на к гиперэкспрессии α-syn.
Рис. 2. Инфекция CVB3 индуцирует образование телец-включений, подобных тельцам Леви, в нейронах.
(A) ICC-изображения клеток WT, α-syn OE dSH-SY5Y и первичных кортикальных нейронов мыши, инфицированных CVB3 (MOI 0,25 или 5) в течение 24 часов. Клетки иммуноокрашивали на CVB3 VP1 (зеленый) и MAP1LC3B (LC3) (красный). (B) ICC-изображения клеток α-syn OE dSH-SY5Y и первичных кортикальных нейронов мыши, инфицированных CVB3 в течение 24 часов. Клетки иммуноокрашивали на CVB3 VP1 (красный) и pSer129 α-syn (зеленый). (C) ICC-изображения клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Наблюдалась совместная локализация CVB3 VP1 (красный) и убиквитина (зеленый). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (D) Просвечивающая электронная микроскопия (ПЭМ) изображения контрольных и инфицированных CVB3 клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов.(E) TEM-изображения клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов. Стрелки указывают на вирусные частицы. (F) ПЭМ-изображения первичных корковых нейронов контрольной группы и мыши, инфицированных CVB3, в течение 24 часов. Стрелки указывают на вирусные частицы. (G) Была проанализирована длина α-syn фибрилл между CVB3-инфицированными клетками WT и α-syn OE dSH-SY5Y.
Клетки иммуноокрашивали на CVB3 VP1 (зеленый) и MAP1LC3B (LC3) (красный). (B) ICC-изображения клеток α-syn OE dSH-SY5Y и первичных кортикальных нейронов мыши, инфицированных CVB3 в течение 24 часов. Клетки иммуноокрашивали на CVB3 VP1 (красный) и pSer129 α-syn (зеленый). (C) ICC-изображения клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Наблюдалась совместная локализация CVB3 VP1 (красный) и убиквитина (зеленый). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (D) Просвечивающая электронная микроскопия (ПЭМ) изображения контрольных и инфицированных CVB3 клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов.(E) TEM-изображения клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов. Стрелки указывают на вирусные частицы. (F) ПЭМ-изображения первичных корковых нейронов контрольной группы и мыши, инфицированных CVB3, в течение 24 часов. Стрелки указывают на вирусные частицы. (G) Была проанализирована длина α-syn фибрилл между CVB3-инфицированными клетками WT и α-syn OE dSH-SY5Y. Стрелки указывают на структуры, подобные α-син фибриллам. * P <0,05, непарный t-критерий. (H) Была проанализирована длина фибрилл α-syn между инфицированными CVB3 WT и первичными нейронами α-syn TG.Стрелки указывают на структуры, подобные α-син фибриллам. ** P <0,01, непарный t-критерий. (I) ПЭМ-изображения контрольных и инфицированных CVB3 (24 ч) клеток WT и α-syn OE dSH-SY5Y. (J) TEM-анализ митохондриальных типов клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 в течение 24 часов. *** P <0,001, однофакторный дисперсионный анализ с тестом множественных сравнений Тьюки.
Стрелки указывают на структуры, подобные α-син фибриллам. * P <0,05, непарный t-критерий. (H) Была проанализирована длина фибрилл α-syn между инфицированными CVB3 WT и первичными нейронами α-syn TG.Стрелки указывают на структуры, подобные α-син фибриллам. ** P <0,01, непарный t-критерий. (I) ПЭМ-изображения контрольных и инфицированных CVB3 (24 ч) клеток WT и α-syn OE dSH-SY5Y. (J) TEM-анализ митохондриальных типов клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 в течение 24 часов. *** P <0,001, однофакторный дисперсионный анализ с тестом множественных сравнений Тьюки.
https://doi.org/10.1371/journal.ppat.1010018.g002
α-Syn регулирует созревание LB-подобных телец включения, индуцированное CVB3
Мы более подробно проанализировали связь между LB-подобными тельцами включения, образованными CVB3 и α-syn.CVB3 индуцировал образование различных типов LB-подобных телец включения с течением времени, что оценивалось по образцам окрашивания LC3. Мы разделили их на четыре стадии на основе картины окрашивания LC3 (рис. 3А). Наблюдали VP1 CVB3, и стадия, когда внутриклеточная морфология LC3 не отличалась от неинфицированных клеток, была определена как стадия 1 (рис. 3A1). Стадия, на которой внутриклеточное расположение LC3 начало претерпевать небольшие изменения, была определена как стадия 2 (рис. 3А2), а стадия, на которой LC3 начала формировать сферу, была определена как стадия 3 (рис. 3А3).Наконец, этап, на котором LC3 сформировал полную сферу с высокой интенсивностью, был определен как этап 4 (рис. 3A4). Эти структуры также колокализуются с pSer129 α-syn по мере развития стадии. Со временем количество телец включения на стадии 4 увеличилось (рис. 3B и 3C), что свидетельствует о созревании LB-подобных телец включения. Интересно, что количество телец включения на стадии 4 было выше в клетках α-syn OE SH-SY5Y и ниже в клетках α-syn KO (по сравнению с контролем) (рис. 3B–3D). Кроме того, в клетках α-syn OE SH-SY5Y LC3 был локализован с тельцами включения с большей интенсивностью, чем в контроле и клетках α-syn KO (рис.
3А). Наблюдали VP1 CVB3, и стадия, когда внутриклеточная морфология LC3 не отличалась от неинфицированных клеток, была определена как стадия 1 (рис. 3A1). Стадия, на которой внутриклеточное расположение LC3 начало претерпевать небольшие изменения, была определена как стадия 2 (рис. 3А2), а стадия, на которой LC3 начала формировать сферу, была определена как стадия 3 (рис. 3А3).Наконец, этап, на котором LC3 сформировал полную сферу с высокой интенсивностью, был определен как этап 4 (рис. 3A4). Эти структуры также колокализуются с pSer129 α-syn по мере развития стадии. Со временем количество телец включения на стадии 4 увеличилось (рис. 3B и 3C), что свидетельствует о созревании LB-подобных телец включения. Интересно, что количество телец включения на стадии 4 было выше в клетках α-syn OE SH-SY5Y и ниже в клетках α-syn KO (по сравнению с контролем) (рис. 3B–3D). Кроме того, в клетках α-syn OE SH-SY5Y LC3 был локализован с тельцами включения с большей интенсивностью, чем в контроле и клетках α-syn KO (рис. 3E и 3F).Тельца включения были более конденсированы, и доля CVB3, присутствующая в тельцах включения, также была увеличена (рис. 3E и 3G-3H), что позволяет предположить, что сверхэкспрессия α-син ускоряет созревание телец включения, образование которых индуцируется CVB3.
3E и 3F).Тельца включения были более конденсированы, и доля CVB3, присутствующая в тельцах включения, также была увеличена (рис. 3E и 3G-3H), что позволяет предположить, что сверхэкспрессия α-син ускоряет созревание телец включения, образование которых индуцируется CVB3.
Рис. 3. α-Syn регулирует созревание телец включения, подобных тельцам Леви, индуцированное CVB3.
(A) Изображения ICC клеток dSH-SY5Y, инфицированных CVB3 (MOI 0,25) в течение 24 часов. Клетки иммуноокрашивали на CVB3 VP1 (зеленый) и MAP1LC3B (LC3) (красный).Более высокое увеличение изображений, заключенных в пронумерованные белые прямоугольники, указывает на стадии тел включения на основе картины окрашивания LC3. Клетки иммуноокрашивали на CVB3 VP1 (зеленый) и pSer129 α-syn (красный). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (B) Анализ изображения ICC соотношения телец включения на каждой стадии в клетках dSH-SY5Y, инфицированных CVB3, в течение 6 и 24 часов. *** P <0,001, * P <0,05, двухфакторный ANOVA тест с тестом множественных сравнений Сидака. (C) Анализ изображения ICC соотношения телец включения на каждой стадии клеток α-syn OE dSH-SY5Y, инфицированных CVB3, в течение 6 и 24 часов.*** P <0,001, двухфакторный анализ ANOVA с тестом множественных сравнений Сидака. (D) Анализ изображения ICC процентного содержания телец включения на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, которые были инфицированы 0,25 MOI CVB3 в течение 24 часов. *** P <0,001, ** P <0,01, * P <0,05, двухфакторный ANOVA тест с тестом множественных сравнений Сидака. (E) ICC-изображения паттернов колокализации VP1 (зеленый) и LC3 (красный) в клетках WT, α-syn OE и α-syn KO dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Изображения являются репрезентативными для независимых экспериментов (n = 3).Масштабная линейка указывает 2 мкм. Синий указывает на Hoechst. (F и G) Анализ изображения ICC интенсивности LC3 и объема телец включения на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Значения получены из трех независимых экспериментов (n = 3).
(C) Анализ изображения ICC соотношения телец включения на каждой стадии клеток α-syn OE dSH-SY5Y, инфицированных CVB3, в течение 6 и 24 часов.*** P <0,001, двухфакторный анализ ANOVA с тестом множественных сравнений Сидака. (D) Анализ изображения ICC процентного содержания телец включения на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, которые были инфицированы 0,25 MOI CVB3 в течение 24 часов. *** P <0,001, ** P <0,01, * P <0,05, двухфакторный ANOVA тест с тестом множественных сравнений Сидака. (E) ICC-изображения паттернов колокализации VP1 (зеленый) и LC3 (красный) в клетках WT, α-syn OE и α-syn KO dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Изображения являются репрезентативными для независимых экспериментов (n = 3).Масштабная линейка указывает 2 мкм. Синий указывает на Hoechst. (F и G) Анализ изображения ICC интенсивности LC3 и объема телец включения на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, инфицированных CVB3 в течение 24 часов. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (H) Анализ интенсивности изображения ICC VP1 в тельцах включения на общую интенсивность VP1 на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов.Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки.
*** P <0,001, ** P <0,01, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (H) Анализ интенсивности изображения ICC VP1 в тельцах включения на общую интенсивность VP1 на каждой стадии в клетках WT, α-syn-OE и α-syn KO dSH-SY5Y, которые были инфицированы CVB3 в течение 24 часов.Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, ** P <0,01, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки.
https://doi.org/10.1371/journal.ppat.1010018.g003
α-Syn регулирует репликацию CVB3 в нейронах
Крупные аутофагосомы, индуцированные CVB3, служат репликационными комплексами вируса [34]. Принимая во внимание, что α-син избыточная экспрессия ускоряется созревание телец включения, образование которых было вызвано CVB3 и что & alpha; син OE DSH-SY5Y клетки отображаются большую интенсивность VP1 в телец включения, чем DSH-SY5Y клеток, мы исследовали, влияет ли α-син репликации CVB3.Репликация CVB3 была увеличена в клетках α-syn OE dSH-SY5Y. Напротив, репликация CVB3 была снижена в клетках α-syn KO dSH-SY5Y. Первичные нейроны от мышей α-syn TG также показали аналогичные результаты (рис. 4A), что согласуется с анализом TEM (рис. 2E). Инфекция вариантом CVB3, коэкспрессирующим EGFP (CVB3-EGFP), приводила к увеличению количества EGFP-позитивных клеток и повышала интенсивность EGFP в клетках α-syn OE SH-SY5Y (рис. 4B–4E). Кроме того, когда клетки SH-SY5Y, сверхэкспрессирующие только mCherry или α-syn-mCherry, были инфицированы CVB3-EGFP, интенсивность EGFP была положительно пропорциональна таковой у mCherry в клетках α-syn-mCherry OE SH-SY5Y, но не в клетках mCherry. только клетки OE SH-SY5Y (фиг. 4F и 4G).Кроме того, когда обе клетки были инфицированы CVB3 в течение 30 часов, цитотоксичность инфекции CVB3 была более серьезной в клетках α-syn OE SH-SY5Y (рис. 4H), что позволяет предположить, что α-syn способствует репликации CVB3 и связанной с ней цитотоксичности.
Напротив, репликация CVB3 была снижена в клетках α-syn KO dSH-SY5Y. Первичные нейроны от мышей α-syn TG также показали аналогичные результаты (рис. 4A), что согласуется с анализом TEM (рис. 2E). Инфекция вариантом CVB3, коэкспрессирующим EGFP (CVB3-EGFP), приводила к увеличению количества EGFP-позитивных клеток и повышала интенсивность EGFP в клетках α-syn OE SH-SY5Y (рис. 4B–4E). Кроме того, когда клетки SH-SY5Y, сверхэкспрессирующие только mCherry или α-syn-mCherry, были инфицированы CVB3-EGFP, интенсивность EGFP была положительно пропорциональна таковой у mCherry в клетках α-syn-mCherry OE SH-SY5Y, но не в клетках mCherry. только клетки OE SH-SY5Y (фиг. 4F и 4G).Кроме того, когда обе клетки были инфицированы CVB3 в течение 30 часов, цитотоксичность инфекции CVB3 была более серьезной в клетках α-syn OE SH-SY5Y (рис. 4H), что позволяет предположить, что α-syn способствует репликации CVB3 и связанной с ней цитотоксичности.
Рис. 4. α-Syn регулирует репликацию CVB3 в нейронах.
(A) Относительные уровни VP1, проанализированные с помощью ПЦР и анализа бляшек, между клетками WT, α-syn OE и α-syn KO dSH-SY5Y, которые были инфицированы CVB3 (MOI 0,25) в течение 24 часов. В случае анализа бляшек каждую клетку инфицировали CVB3 (MOI 1) в течение 30 часов.Относительные уровни VP1, проанализированные с помощью ПЦР, между первичными кортикальными нейронами мышей WT и α-syn TG, которые были инфицированы CVB3 (MOI 5) в течение 24 часов. Значения получены из шести или трех независимых экспериментов (n = 6 или 3). *** P <0,001, ** P <0,01, * P <0,05, однофакторный дисперсионный анализ с тестом множественных сравнений Тьюки и непарным t-тестом. (B) Изображения флуоресцентной микроскопии клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы вариантом CVB3 0,6 MOI, коэкспрессирующим EGFP (CVB3-EGFP), в течение 24 часов.Масштабная линейка указывает 50 мкм. (C, D и E) Проточный цитометрический анализ. Количество EGFP-позитивных клеток (D) и интенсивность (E) анализировали с использованием клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения получены из трех или четырех независимых экспериментов (n = 3 или 4). * P <0,05, непарный t-критерий.
В случае анализа бляшек каждую клетку инфицировали CVB3 (MOI 1) в течение 30 часов.Относительные уровни VP1, проанализированные с помощью ПЦР, между первичными кортикальными нейронами мышей WT и α-syn TG, которые были инфицированы CVB3 (MOI 5) в течение 24 часов. Значения получены из шести или трех независимых экспериментов (n = 6 или 3). *** P <0,001, ** P <0,01, * P <0,05, однофакторный дисперсионный анализ с тестом множественных сравнений Тьюки и непарным t-тестом. (B) Изображения флуоресцентной микроскопии клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы вариантом CVB3 0,6 MOI, коэкспрессирующим EGFP (CVB3-EGFP), в течение 24 часов.Масштабная линейка указывает 50 мкм. (C, D и E) Проточный цитометрический анализ. Количество EGFP-позитивных клеток (D) и интенсивность (E) анализировали с использованием клеток WT и α-syn OE dSH-SY5Y, которые были инфицированы CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения получены из трех или четырех независимых экспериментов (n = 3 или 4). * P <0,05, непарный t-критерий. (F) Изображения флуоресцентной микроскопии клеток mCherry и α-syn-mCherry OE dSH-SY5Y, инфицированных CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения и значения получены из трех независимых экспериментов (n = 3).Масштабная линейка указывает 50 мкм. (G) Линейный регрессионный анализ интенсивности mCherry и EGFP клеток mCherry и α-syn-mCherry OE dSH-SY5Y, инфицированных CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения являются репрезентативными для трех независимых экспериментов. (H) Анализ цитотоксичности клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 (MOI 0,25) в течение 30 ч с помощью анализа лактатдегидрогеназы. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, непарный t-критерий.
(F) Изображения флуоресцентной микроскопии клеток mCherry и α-syn-mCherry OE dSH-SY5Y, инфицированных CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения и значения получены из трех независимых экспериментов (n = 3).Масштабная линейка указывает 50 мкм. (G) Линейный регрессионный анализ интенсивности mCherry и EGFP клеток mCherry и α-syn-mCherry OE dSH-SY5Y, инфицированных CVB3-EGFP (MOI 0,6) в течение 24 часов. Значения являются репрезентативными для трех независимых экспериментов. (H) Анализ цитотоксичности клеток WT и α-syn OE dSH-SY5Y, инфицированных CVB3 (MOI 0,25) в течение 30 ч с помощью анализа лактатдегидрогеназы. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, непарный t-критерий.
https://дои.org/10.1371/journal.ppat.1010018.g004
CVB3 и α-syn по-разному регулируют аутофагическую активность
CVB3 ингибирует слияние аутофагосом с лизосомами и использует аутофагосомы в качестве репликационных комплексов [34]. Чтобы подтвердить это, мы отслеживали аутофагическую активность [35, 36]. Уровни LC3II повышались при инфицировании CVB3. Однако он не увеличивался дальше при лечении бафиломицином А1 (BafA1) (рис. 5А), что свидетельствует о том, что CVB3 ингибирует позднюю стадию аутофагического процесса.Это также было подтверждено значительным снижением лизосом, по оценке окрашивания LysoTracker, в CVB3-инфицированных клетках dSH-SY5Y (рис. 5B). Кроме того, уровни LC3II повышались, а обработка BafA1 дополнительно увеличивала уровни LC3II в клетках α-syn OE dSH-SY5Y (рис. 5C). Уровни р62 были снижены (рис. 5D), что свидетельствует о том, что сверхэкспрессия α-син приводит к усилению потока аутофагии, что согласуется с результатами предыдущего исследования [37]. Далее мы проанализировали информацию из базы данных с открытым исходным кодом.Сверхэкспрессия α-syn человека с использованием лентивирусного вектора в нейронах среднего мозга мыши показала, что кластер транскриптома характеризовался «положительной регуляцией макроаутофагии» (GO:0016239) в анализе генной онтологии (GO) по сравнению с контролем (GSE70368) (S3 Инжир). Кроме того, «созревание аутофагосом» (GO:0097352) в индуцированных плюрипотентных стволовых клетках (ИПСК) семейства α-syn ( SNCA ) с тройным повторением (GSE30792) и «организация лизосом» (GO:0007040) в ткани стриатума мыши α-syn TG мышей человека (GSE116010) также были выше, чем в контроле (S3 Fig).Эти выводы подтвердили наши данные. По сравнению с контролем, инфекция CVB3 дополнительно повышала уровни LC3 II, а обработка BafA1 индуцировала аналогичные результаты в клетках α-syn OE dSH-SY5Y (рис. 5E и 5F). Эти результаты свидетельствуют о том, что сверхэкспрессия α-syn способствует аутофагическому потоку и ускоряет образование аутофагосом, которые обеспечивают больше центров репликации для CVB3.
Кроме того, «созревание аутофагосом» (GO:0097352) в индуцированных плюрипотентных стволовых клетках (ИПСК) семейства α-syn ( SNCA ) с тройным повторением (GSE30792) и «организация лизосом» (GO:0007040) в ткани стриатума мыши α-syn TG мышей человека (GSE116010) также были выше, чем в контроле (S3 Fig).Эти выводы подтвердили наши данные. По сравнению с контролем, инфекция CVB3 дополнительно повышала уровни LC3 II, а обработка BafA1 индуцировала аналогичные результаты в клетках α-syn OE dSH-SY5Y (рис. 5E и 5F). Эти результаты свидетельствуют о том, что сверхэкспрессия α-syn способствует аутофагическому потоку и ускоряет образование аутофагосом, которые обеспечивают больше центров репликации для CVB3.
Рис. 5. CVB3 и α-syn по-разному регулируют аутофагическую активность.
(A) Контрольные образцы и образцы, инфицированные CVB3 (0.25 MOI в течение 24 ч) DSH-SY5Y клетки, обработанные диметилсульфоксиде (ДМСО) или 50 нМ bafilomycin A1 (BafA1) лизировали и Вестерн-блоттинг проводили с использованием LC3 антитела.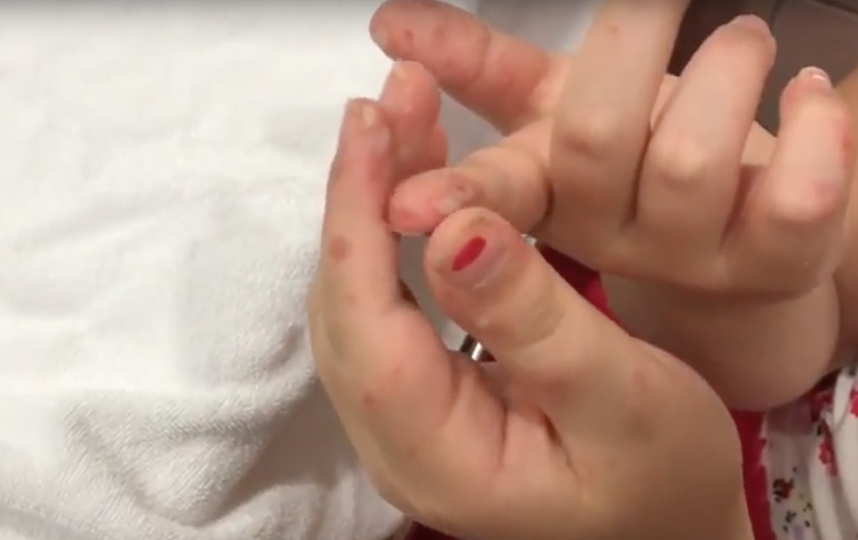 Уровни белка количественно определяли с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). ** P <0,01, односторонний ANOVA тест с несколько тестом сравнения Тьюки. (Б) ICC изображения DSH-SY5Y клеток, инфицированных CVB3 (MOI 0,25) в течение 24 ч. Флуоресцентная был замечен с использованием Lysotracker. Значения получены из трех независимых экспериментов (n = 3).Масштабная линейка указывает 10 мкм. *** P <0,001, непарный t-критерий. (C) Относительный уровень экспрессии p62 между клетками WT и α-syn OE dSH-SY5Y. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, непарный t-критерий. (D) Относительный уровень экспрессии LC3 между клетками WT и α-syn OE dSH-SY5Y, обработанными ДМСО или 50 нМ BafA1 в течение 24 часов. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3).*** P <0,001, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки.
Уровни белка количественно определяли с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). ** P <0,01, односторонний ANOVA тест с несколько тестом сравнения Тьюки. (Б) ICC изображения DSH-SY5Y клеток, инфицированных CVB3 (MOI 0,25) в течение 24 ч. Флуоресцентная был замечен с использованием Lysotracker. Значения получены из трех независимых экспериментов (n = 3).Масштабная линейка указывает 10 мкм. *** P <0,001, непарный t-критерий. (C) Относительный уровень экспрессии p62 между клетками WT и α-syn OE dSH-SY5Y. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, непарный t-критерий. (D) Относительный уровень экспрессии LC3 между клетками WT и α-syn OE dSH-SY5Y, обработанными ДМСО или 50 нМ BafA1 в течение 24 часов. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3).*** P <0,001, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки. (E и F) Относительные уровни экспрессии LC3 между клетками WT (контроль и CVB3) и α-syn OE (контроль и CVB3) dSH-SY5Y с ДМСО или 50 нМ BafA1. Клетки инфицировали CVB3 0,25 MOI в течение 24 часов. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки.
(E и F) Относительные уровни экспрессии LC3 между клетками WT (контроль и CVB3) и α-syn OE (контроль и CVB3) dSH-SY5Y с ДМСО или 50 нМ BafA1. Клетки инфицировали CVB3 0,25 MOI в течение 24 часов. Уровни белка определяли количественно с помощью денситометрии. Значения получены из трех независимых экспериментов (n = 3). *** P <0,001, * P <0,05, однофакторный ANOVA тест с тестом множественных сравнений Тьюки.
https://дои.org/10.1371/journal.ppat.1010018.g005
α-Syn регулирует репликацию CVB3 в мозге мышей
Чтобы подтвердить, взаимодействует ли CVB3 также с α-syn в головном мозге, CVB3 вводили мышам внутрибрюшинно. Экспрессия α-Syn у мышей TG управлялась промотором NSE и была в 1,5 раза выше, чем у мышей WT (фиг. S4A). В наших экспериментальных условиях у мышей TG без инфекции CVB3 не наблюдалось никаких клинических или гистологических аномалий. На 7-й день после заражения (PI) обнаружение CVB3 было более выраженным в мозге мышей α-syn TG по сравнению с таковым у мышей WT (рис. 6A).Данные in vivo соответствовали данным in vitro . Иммуногистохимия (ИГХ) головного мозга мышей WT выявила присутствие CVB3 в обонятельной области, передней поясной извилине, латеральном септальном ядре, области гиппокампа, фимбриях, корково-спинномозговом пути и гипоталамусе, в основном вдоль желудочков, которые были колокализованы с микроглией. маркер, Иба-1. Кроме того, CVB3 был обнаружен в гиппокампе, латеральном таламусе и среднем мозге и локализован вместе с нейрональным маркером Tuj-1.CVB3 не локализовался совместно с маркером астроцитов, GFAP (рис. 6B и 6C), что свидетельствует о том, что CVB3 был обнаружен в головном мозге и инфицировал микроглию и нейроны, но не астроциты.
6A).Данные in vivo соответствовали данным in vitro . Иммуногистохимия (ИГХ) головного мозга мышей WT выявила присутствие CVB3 в обонятельной области, передней поясной извилине, латеральном септальном ядре, области гиппокампа, фимбриях, корково-спинномозговом пути и гипоталамусе, в основном вдоль желудочков, которые были колокализованы с микроглией. маркер, Иба-1. Кроме того, CVB3 был обнаружен в гиппокампе, латеральном таламусе и среднем мозге и локализован вместе с нейрональным маркером Tuj-1.CVB3 не локализовался совместно с маркером астроцитов, GFAP (рис. 6B и 6C), что свидетельствует о том, что CVB3 был обнаружен в головном мозге и инфицировал микроглию и нейроны, но не астроциты.
Рис. 6. α-Syn регулирует репликацию CVB3 в мозге мышей.
(A) Относительные уровни VP1, проанализированные с помощью ПЦР, в мозге мышей WT (n = 8) и α-syn TG (n = 14) и анализ бляшек между WT (n = 7) и α-syn Мозг мышей TG (n = 4) на 7-й день после заражения (PI). Мышей заражали внутрибрюшинной инъекцией 1.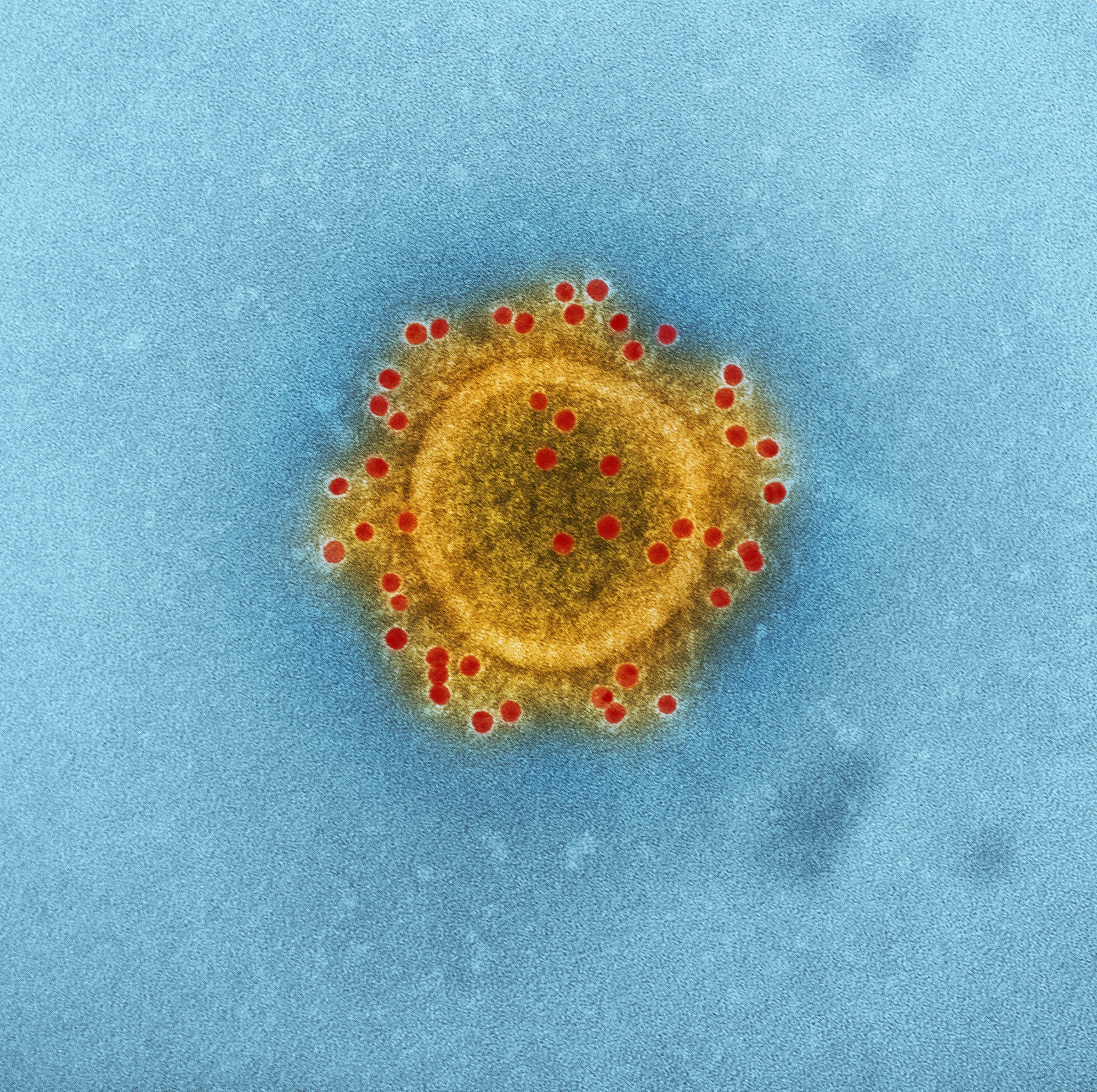 0 × 10 6 БОЕ CVB3. * P <0,05, непарный t-критерий. (B) Изображения иммуногистохимии (IHC), показывающие локализацию VP1-положительных клеток в областях мозга мыши на 4-й день PI. Микроглия и нейроны обозначены зелеными и красными звездочками соответственно. (Сокращения: области мозга мыши: CTX-кора, MOB-основная обонятельная луковица, AON-переднее обонятельное ядро, LSX-латеральный перегородочный комплекс, HY-гипоталамус, ACA-передняя поясная извилина, fxs-система свода, MO-соматомоторная зона, HCF -гиппокампальная формация, TH-таламус, CP-каудопутамен, Cst-корково-спинномозговой путь, RSP-ретросплениальная область, PRT-претектальная область, MRN-ретикулярное ядро среднего мозга, SN-substantia nigra).(C) ИГХ-изображения мозга мышей, инфицированных CVB3, на 4-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Наблюдалась совместная локализация CVB3 VP1 (зеленый) с Iba-1 (красный), Tuj-1 (красный) и GFAP (красный). Масштабная линейка указывает 10 мкм.
0 × 10 6 БОЕ CVB3. * P <0,05, непарный t-критерий. (B) Изображения иммуногистохимии (IHC), показывающие локализацию VP1-положительных клеток в областях мозга мыши на 4-й день PI. Микроглия и нейроны обозначены зелеными и красными звездочками соответственно. (Сокращения: области мозга мыши: CTX-кора, MOB-основная обонятельная луковица, AON-переднее обонятельное ядро, LSX-латеральный перегородочный комплекс, HY-гипоталамус, ACA-передняя поясная извилина, fxs-система свода, MO-соматомоторная зона, HCF -гиппокампальная формация, TH-таламус, CP-каудопутамен, Cst-корково-спинномозговой путь, RSP-ретросплениальная область, PRT-претектальная область, MRN-ретикулярное ядро среднего мозга, SN-substantia nigra).(C) ИГХ-изображения мозга мышей, инфицированных CVB3, на 4-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Наблюдалась совместная локализация CVB3 VP1 (зеленый) с Iba-1 (красный), Tuj-1 (красный) и GFAP (красный). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (D и E) Временные ряды (на 4, 7 и 28 день PI) анатомическая диаграмма, иллюстрирующая характер инфекции CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Положительные плотности клеток Tuj-1 (D) и Iba-1 (E), которые были локализованы совместно с VP1 у мышей дикого типа и α-syn TG, инфицированных CVB3, выражены соответствующим цветом указанного числа в зависимости от анатомической структуры.(F) ИГХ-изображения головного мозга контрольных и инфицированных CVB3 мышей на 28-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Стрелки указывают на расщепленные каспаза-3 (с-каспаза-3) положительные клетки. (Сокращения структуры мозга мыши: Hp: гиппокамп, SN: черная субстанция). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (G) Вестерн-блоттинг проводили с использованием контрольных и инфицированных CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3) WT (на 28-й день PI) и α-syn TG (на 7-й день PI) образцов мозга мышей.
Синий указывает на Hoechst. (D и E) Временные ряды (на 4, 7 и 28 день PI) анатомическая диаграмма, иллюстрирующая характер инфекции CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Положительные плотности клеток Tuj-1 (D) и Iba-1 (E), которые были локализованы совместно с VP1 у мышей дикого типа и α-syn TG, инфицированных CVB3, выражены соответствующим цветом указанного числа в зависимости от анатомической структуры.(F) ИГХ-изображения головного мозга контрольных и инфицированных CVB3 мышей на 28-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Стрелки указывают на расщепленные каспаза-3 (с-каспаза-3) положительные клетки. (Сокращения структуры мозга мыши: Hp: гиппокамп, SN: черная субстанция). Масштабная линейка указывает 10 мкм. Синий указывает на Hoechst. (G) Вестерн-блоттинг проводили с использованием контрольных и инфицированных CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3) WT (на 28-й день PI) и α-syn TG (на 7-й день PI) образцов мозга мышей. Уровни белка определяли количественно с помощью денситометрии. *** P <0,001, * P <0,05, непарный t-критерий. (H) ИГХ-изображения мозга мышей, инфицированных CVB3, на 28-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Наблюдалась совместная локализация CVB3 VP1 (зеленый) с тирозингидроксилазой (TH) (красный). Масштабная линейка указывает 20 мкм. Синий указывает на Hoechst. (I) TH-положительный диаминобензидин (DAB) изображение головного мозга мышей, инфицированных CVB3 и контрольной группы, на 28-й день PI (WT) и на 7-й день PI (α-syn TG). Было проанализировано количество ТГ-позитивных клеток в компактной части черной субстанции (SNpc) между контрольными и инфицированными CVB мышами.Масштабная линейка указывает 100 мкм. *** P <0,001, ** P <0,01, непарный t-критерий. (J) Вестерн-блоттинг проводили с использованием контрольных и инфицированных CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3) мышей WT (на 28-й день PI) и α-syn TG (на 7-й день PI) мышей.
Уровни белка определяли количественно с помощью денситометрии. *** P <0,001, * P <0,05, непарный t-критерий. (H) ИГХ-изображения мозга мышей, инфицированных CVB3, на 28-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Наблюдалась совместная локализация CVB3 VP1 (зеленый) с тирозингидроксилазой (TH) (красный). Масштабная линейка указывает 20 мкм. Синий указывает на Hoechst. (I) TH-положительный диаминобензидин (DAB) изображение головного мозга мышей, инфицированных CVB3 и контрольной группы, на 28-й день PI (WT) и на 7-й день PI (α-syn TG). Было проанализировано количество ТГ-позитивных клеток в компактной части черной субстанции (SNpc) между контрольными и инфицированными CVB мышами.Масштабная линейка указывает 100 мкм. *** P <0,001, ** P <0,01, непарный t-критерий. (J) Вестерн-блоттинг проводили с использованием контрольных и инфицированных CVB3 (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3) мышей WT (на 28-й день PI) и α-syn TG (на 7-й день PI) мышей. Уровни белка определяли количественно с помощью денситометрии. * P <0,05, непарный t-критерий. (K) Кривые выживаемости Каплана-Мейера и кривая потери массы тела (внутрибрюшинная инъекция 1,0 × 10 6 БОЕ CVB3) контрольных, CVB-инфицированных мышей WT и α-syn TG. *** Р < 0.001, логарифмический критерий (анализ Мантеля-Кокса) (анализ кривой выживания). *** P <0,001, двухфакторный дисперсионный анализ (анализ кривой потери веса). Рисунок 6B, 6D и 6E был изменен после загрузки изображения мозга из открытого источника (http://labs.gaidi.ca/mouse-brain-atlas).
Уровни белка определяли количественно с помощью денситометрии. * P <0,05, непарный t-критерий. (K) Кривые выживаемости Каплана-Мейера и кривая потери массы тела (внутрибрюшинная инъекция 1,0 × 10 6 БОЕ CVB3) контрольных, CVB-инфицированных мышей WT и α-syn TG. *** Р < 0.001, логарифмический критерий (анализ Мантеля-Кокса) (анализ кривой выживания). *** P <0,001, двухфакторный дисперсионный анализ (анализ кривой потери веса). Рисунок 6B, 6D и 6E был изменен после загрузки изображения мозга из открытого источника (http://labs.gaidi.ca/mouse-brain-atlas).
https://doi.org/10.1371/journal.ppat.1010018.g006
Кроме того, CVB3 наблюдали в нейронах, расположенных в гиппокампе, латеральном таламусе и среднем мозге на 4-й день PI. На 7-й день PI количество нейронов, инфицированных CVB3, было увеличено по сравнению с таковым у мышей на 4-й день PI (рис. 6D и S4B).На 28-й день PI количество нейронов среднего мозга, инфицированных CVB3, было явно увеличено, тогда как количество нейронов, расположенных в других областях, инфицированных CVB3, было относительно уменьшено (рис. 6D). Паттерн инфицированных нейронов был более ускоренным у мышей α-syn TG, чем у мышей WT. На 4-й день PI количество нейронов, инфицированных CVB3, существенно не отличалось между мышами дикого типа и мышами α-syn TG, но в гиппокампе мышей α-syn TG наблюдалось относительно больше нейронов, инфицированных CVB3, по сравнению с таковым у мышей дикого типа. мыши (рис. 6D).На 7-й день PI картина инфицированных нейронов у мышей TG была сопоставима с таковой у мышей WT на 28-й день PI (рис. 6D и S4B). Количество микроглии, инфицированной CVB3, увеличилось на 7-й день PI (рис. 6E и S4C). На 28-й день PI количество микроглии, инфицированной CVB3, было таким же, как и на 7-й день PI (рис. 6E и S4C). Количество микроглии, инфицированной CVB3, также было выше у мышей TG, чем у мышей WT в течение того же периода (рис. 6E и S4C). Кроме того, при заражении CVB3 наблюдали активацию микроглии (рис. S5).
6D). Паттерн инфицированных нейронов был более ускоренным у мышей α-syn TG, чем у мышей WT. На 4-й день PI количество нейронов, инфицированных CVB3, существенно не отличалось между мышами дикого типа и мышами α-syn TG, но в гиппокампе мышей α-syn TG наблюдалось относительно больше нейронов, инфицированных CVB3, по сравнению с таковым у мышей дикого типа. мыши (рис. 6D).На 7-й день PI картина инфицированных нейронов у мышей TG была сопоставима с таковой у мышей WT на 28-й день PI (рис. 6D и S4B). Количество микроглии, инфицированной CVB3, увеличилось на 7-й день PI (рис. 6E и S4C). На 28-й день PI количество микроглии, инфицированной CVB3, было таким же, как и на 7-й день PI (рис. 6E и S4C). Количество микроглии, инфицированной CVB3, также было выше у мышей TG, чем у мышей WT в течение того же периода (рис. 6E и S4C). Кроме того, при заражении CVB3 наблюдали активацию микроглии (рис. S5).
На 28-й день PI экспрессия расщепленной каспазы 3, маркера апоптоза, локально наблюдалась по всему мозгу мышей дикого типа (рис. 6F и S6). Это наблюдение было также очевидно у мышей TG на 7-й день PI, что было сравнимо с наблюдением у мышей WT на 28-й день PI (рис. 6G). Кроме того, дофаминергические нейроны, расположенные в черной субстанции, были инфицированы CVB3 (рис. 6H). Количество клеток, положительных по тирозингидроксилазе (TH), было немного снижено в черной субстанции мышей, инфицированных CVB3, на 28-й день ИП по сравнению с контролем, что также было сопоставимо с таковым у мышей TG на 7-й день ИП (рис. 6I и 6J).
6F и S6). Это наблюдение было также очевидно у мышей TG на 7-й день PI, что было сравнимо с наблюдением у мышей WT на 28-й день PI (рис. 6G). Кроме того, дофаминергические нейроны, расположенные в черной субстанции, были инфицированы CVB3 (рис. 6H). Количество клеток, положительных по тирозингидроксилазе (TH), было немного снижено в черной субстанции мышей, инфицированных CVB3, на 28-й день ИП по сравнению с контролем, что также было сопоставимо с таковым у мышей TG на 7-й день ИП (рис. 6I и 6J).
Затем мы наблюдали за выживаемостью как мышей дикого типа, так и мышей α-syn TG. В отсутствие инфекции CVB3 не было различий в прибавке веса между мышами WT и α-syn TG, и обе группы мышей выжили в наших экспериментальных условиях. Однако после заражения CVB3 выживаемость мышей α-syn TG была ниже, чем у мышей дикого типа. Потеря веса была более значительной у мышей α-syn TG. Эти данные свидетельствуют о том, что мыши α-syn TG были более восприимчивы к инфекции CVB3 (рис. 6K). CVB3 — кардиотропный вирус, вызывающий миокардит [16]. Поэтому мы отслеживали повреждение миокарда и репликацию CVB3 в сердце. Повреждения сердца и уровни VP1 в миокарде обеих групп мышей были сходными (S7A и S7B на фиг.1). Кроме того, CVB3 вызывают обширное повреждение ткани поджелудочной железы в экспериментальной модели животных [38, 39]. Гистологическое исследование поджелудочной железы с окрашиванием гематоксилином и эозином показало повреждение ткани поджелудочной железы с воспалением. Однако никакой разницы между обеими группами мышей не наблюдалось (фиг. S7C). Экспрессия эндогенного α-сина в сердце и поджелудочной железе была намного ниже, чем в мозге, и не было различий в экспрессии эндогенного α-сина в сердце и поджелудочной железе как у мышей WT, так и у мышей TG, поскольку сверхэкспрессия α-сина контролировалась. промотором NSE (фиг. S7D).Эти результаты показали, что разница в выживаемости обеих групп мышей может быть не связана с повреждением сердца или поджелудочной железы. Гистологические исследования сердца, печени и селезенки также не показали различий между мышами WT и α-syn TG (рис. S7C). Эти результаты свидетельствуют о том, что инфекция CVB3 в головном мозге мыши вызывает гибель нейронов, включая потерю дофаминергических нейронов, расположенных в черной субстанции, и что α-syn ускоряет репликацию CVB3 и CVB3-индуцированной гибели нейронов.
S7C). Эти результаты свидетельствуют о том, что инфекция CVB3 в головном мозге мыши вызывает гибель нейронов, включая потерю дофаминергических нейронов, расположенных в черной субстанции, и что α-syn ускоряет репликацию CVB3 и CVB3-индуцированной гибели нейронов.
CVB3 индуцирует образование включений α-син в мозге мышей
Затем мы исследовали совместную локализацию α-syn и CVB3 VP1 в головном мозге мышей, инфицированных CVB3. Мы не наблюдали совместной локализации α-syn и CVB3 VP1 в мозге мышей WT или α-syn TG. Вместо этого на 7-й день PI несколько нейронов, содержащих накопление α-син в теле клетки, наблюдались в межножковом ядре мышей WT после инфекции CVB3, и интенсивность α-син была ниже, чем у контроля (рис. 7A).Однако скопления α-син в теле клетки не локализовались совместно с α-сином pSer129. У мышей α-syn TG в той же области наблюдалось больше нейронов, содержащих накопление α-syn в теле клетки (рис. 7А), хотя они также не колокализовались с pSer129 α-syn. Мы провели вестерн-блоттинг для обнаружения pSer129 α-syn. На 28-й день PI у мышей WT был обнаружен низкий уровень pSer 129 α-syn, и наблюдалось снижение экспрессии α-syn (фиг. 7B). Сходные результаты наблюдались у мышей α-syn TG на 7-й день PI, что свидетельствует о том, что накопление α-syn в теле клетки индуцируется инфекцией CVB3 и ускоряется в ответ на сверхэкспрессию α-syn (рис. 7C).
Мы провели вестерн-блоттинг для обнаружения pSer129 α-syn. На 28-й день PI у мышей WT был обнаружен низкий уровень pSer 129 α-syn, и наблюдалось снижение экспрессии α-syn (фиг. 7B). Сходные результаты наблюдались у мышей α-syn TG на 7-й день PI, что свидетельствует о том, что накопление α-syn в теле клетки индуцируется инфекцией CVB3 и ускоряется в ответ на сверхэкспрессию α-syn (рис. 7C).
Рис. 7. CVB3 индуцирует включения α-син в мозге мышей.
(A) ИГХ-изображения контрольного, CVB3-инфицированного мозга мышей WT и α-syn TG в указанные дни PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Масштабная линейка указывает 20 мкм. Синий указывает на Hoechst. (Сокращения областей мозга мыши: CTX-кора, HCF-образование гиппокампа, MB-средний мозг, TH-thalamus, SN-substantia nigra, IPN-межножковое ядро). (B и C) Вестерн-блоттинг проводили с использованием контроля, CVB3 (в/б инъекция 1.0 × 10 6 БОЕ CVB3) инфицированных образцов мозга мышей WT и α-syn TG в указанные дни PI. Уровни белка определяли количественно с помощью денситометрии. *** P <0,001, ** P <0,01, * P <0,05, непарный t-критерий. Рис. 7А был изменен после загрузки изображения мозга из открытого источника (http://labs.gaidi.ca/mouse-brain-atlas).
https://doi.org/10.1371/journal.ppat.1010018.g007
Обсуждение
Несмотря на обширные исследования, механизм, лежащий в основе патогенеза БП, остается неясным.Предполагается, что как генетические факторы, так и факторы окружающей среды, а также их взаимодействие вносят свой вклад в патогенез БП [40]. LBs являются основными патологическими признаками БП. Кроме того, патология Леви прогрессивно вовлекает больше областей нервной системы по мере прогрессирования заболевания и проявляется до появления двигательных симптомов при БП [8]. Соответственно, важно определить факторы, которые инициируют патологию Леви, чтобы понять патогенез БП. Предполагается, что несколько факторов вызывают патологию Леви.Несколько факторов окружающей среды, которые вносят свой вклад, включают патогены, такие как вирус гриппа, загрязнители окружающей среды, такие как пестициды, тяжелые металлы и травмы головы [41]. В частности, вирусная инфекция долгое время рассматривалась как фактор риска нейродегенеративных заболеваний [13]. Настоящие данные демонстрируют, что инфекция CVB3 и экспрессия α-syn взаимно влияют друг на друга.
В частности, вирусная инфекция долгое время рассматривалась как фактор риска нейродегенеративных заболеваний [13]. Настоящие данные демонстрируют, что инфекция CVB3 и экспрессия α-syn взаимно влияют друг на друга.
Что касается влияния CVB3 на α-syn, инфекция CVB3 индуцирует очень большие структуры, связанные с аутофагией, диаметром от 10 до 20 мкм в нейронах.Эти структуры локализуются совместно с LC3 и имеют сходную морфологию с мегафагосомами, наблюдаемыми в ацинарных клетках поджелудочной железы [28]. Они также колокализованы с α-syn, pSer129 α-syn и убиквитином, что указывает на сходство с LB. ЛБ демонстрируют значительное морфологическое разнообразие и гетерогенны по своей форме, биохимическому составу и организации [30]. При исследовании головного мозга пациентов с БП с помощью микроскопии сверхвысокого разрешения на основе истощения стимулированной эмиссии (STED) были обнаружены ЛБ со скученными органеллами и липидными мембранами.Это вызвало предположение, что α-syn может модулировать компартментализацию и функцию мембран и органелл в пораженных LB клетках [31]. Недавний отчет продемонстрировал образование филаментоподобных структур, сопровождающееся секвестрацией липидов, органелл и эндомембранных структур, с использованием модели фибрилляции α-син, основанной на посевах, которая повторяет особенности LB, наблюдаемые в мозге пациентов с БП. 42]. Точно так же наши результаты ПЭМ также показали, что инфекция CVB3 приводила к скоплению нескольких органелл в околоядерном пространстве нейронов.Переполненные органеллы содержали поврежденные митохондрии и множество фибриллярных структур, окружающих органеллы. Количество фибриллярных структур в клетках α-syn OE dSH-SY5Y было выше, чем в клетках dSH-SY5Y. Эти структуры в клетках α-syn OE dSH-SY5Y также были длиннее, чем в клетках dSH-SY5Y. Эти структуры практически не обнаруживались в клетках α-syn KO. Фибриллы α-син имеют диаметр 20 нм in vitro [43]. Эти наблюдения позволили предположить, что эти фибриллярные структуры могут быть α-син фибриллами, хотя мы не смогли подтвердить это с помощью иммуно-ЭМ-анализа.
Недавний отчет продемонстрировал образование филаментоподобных структур, сопровождающееся секвестрацией липидов, органелл и эндомембранных структур, с использованием модели фибрилляции α-син, основанной на посевах, которая повторяет особенности LB, наблюдаемые в мозге пациентов с БП. 42]. Точно так же наши результаты ПЭМ также показали, что инфекция CVB3 приводила к скоплению нескольких органелл в околоядерном пространстве нейронов.Переполненные органеллы содержали поврежденные митохондрии и множество фибриллярных структур, окружающих органеллы. Количество фибриллярных структур в клетках α-syn OE dSH-SY5Y было выше, чем в клетках dSH-SY5Y. Эти структуры в клетках α-syn OE dSH-SY5Y также были длиннее, чем в клетках dSH-SY5Y. Эти структуры практически не обнаруживались в клетках α-syn KO. Фибриллы α-син имеют диаметр 20 нм in vitro [43]. Эти наблюдения позволили предположить, что эти фибриллярные структуры могут быть α-син фибриллами, хотя мы не смогли подтвердить это с помощью иммуно-ЭМ-анализа.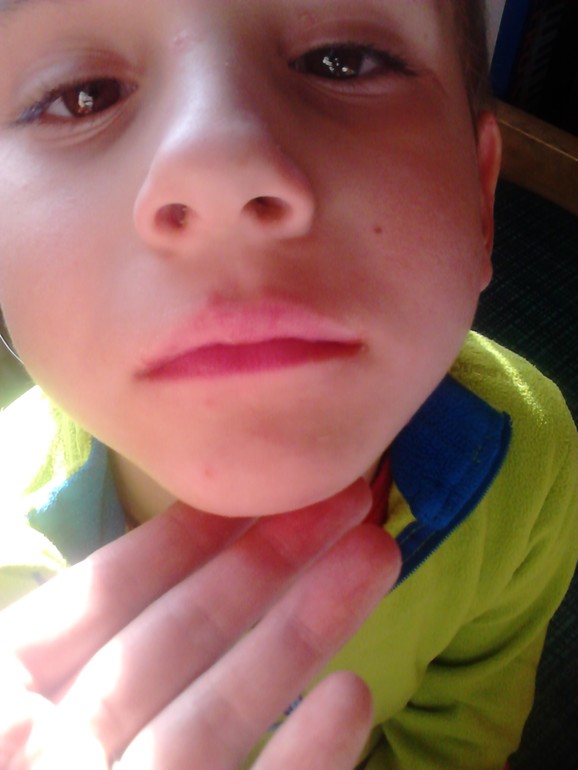 Кроме того, формирование ЛБ включает несколько стадий [44, 45]. CVB3 индуцирует различные типы LB-подобных телец включения с течением времени, что может отражать созревание телец включения. В клетках α-syn OE созревание этих телец включения было ускорено, и они были более конденсированы, что свидетельствует о том, что α-syn может регулировать созревание телец включения в качестве основного компонента. Кроме того, митохондриальное повреждение индуцировалось CVB3, которого было больше в клетках α-syn OE dSH-SY5Y. Наблюдения частично подтверждаются предыдущими исследованиями, демонстрирующими, что α-syn локализуется в митохондриях, а OE-клетки α-syn обнаруживают митохондриальную дисфункцию [46-48].Мы продемонстрировали, что CVB3 ингибирует позднюю стадию аутофагии в клетках dSH-SY5Y, что согласуется с результатами предыдущего исследования [28]. Хорошо известно, что α-син является субстратом для аутофагической деградации [49–51], хотя сообщалось и о других механизмах, участвующих в деградации α-син [52].
Кроме того, формирование ЛБ включает несколько стадий [44, 45]. CVB3 индуцирует различные типы LB-подобных телец включения с течением времени, что может отражать созревание телец включения. В клетках α-syn OE созревание этих телец включения было ускорено, и они были более конденсированы, что свидетельствует о том, что α-syn может регулировать созревание телец включения в качестве основного компонента. Кроме того, митохондриальное повреждение индуцировалось CVB3, которого было больше в клетках α-syn OE dSH-SY5Y. Наблюдения частично подтверждаются предыдущими исследованиями, демонстрирующими, что α-syn локализуется в митохондриях, а OE-клетки α-syn обнаруживают митохондриальную дисфункцию [46-48].Мы продемонстрировали, что CVB3 ингибирует позднюю стадию аутофагии в клетках dSH-SY5Y, что согласуется с результатами предыдущего исследования [28]. Хорошо известно, что α-син является субстратом для аутофагической деградации [49–51], хотя сообщалось и о других механизмах, участвующих в деградации α-син [52]. Сообщалось, что дисфункциональная поздняя стадия аутофагии связана с накоплением α-син [53]. Соответственно, CVB3 может использовать механизм аутофагии, чтобы индуцировать образование LB-подобных включений, связанных с α-syn.
Сообщалось, что дисфункциональная поздняя стадия аутофагии связана с накоплением α-син [53]. Соответственно, CVB3 может использовать механизм аутофагии, чтобы индуцировать образование LB-подобных включений, связанных с α-syn.
Хотя CVB3 индуцировал образование больших телец включения, содержащих α-син, его экспрессия была снижена. Мы подтвердили это, используя модельные системы in vitro и in vivo и данные из открытых источников по сердцам мышей, инфицированных CVB3. В частности, соседние клетки могут быть более затронуты. Это может быть не связано с цитотоксичностью или феноменом отключения хозяина при заражении CVB3. Тем не менее полностью игнорировать их влияние нельзя. Дифференциальная регуляция экспрессии α-syn требует дальнейшего изучения.В предыдущем отчете было показано, что ВЗН индуцирует экспрессию α-син, и α-син был предложен в качестве фактора рестрикции вируса [21]. Мы также наблюдали, что обработка полиИК увеличивала экспрессию α-син в клетках dSH-SY5Y, что свидетельствует о том, что экспрессия α-син может регулироваться вирусной инфекцией. Однако снижение экспрессии α-syn может быть специфичным для CVB3. Интересно, что в головном мозге пациентов с БП было описано снижение уровня мРНК α-син [54, 55]. Эти результаты являются спорными из-за нескольких технических проблем, включая методы выборки и нормализации.Наш анализ открытых источников данных о пациентах с БП также подтвердил это. Следовательно, инфицирование CVB3 у пациентов может свидетельствовать о начале БП.
Однако снижение экспрессии α-syn может быть специфичным для CVB3. Интересно, что в головном мозге пациентов с БП было описано снижение уровня мРНК α-син [54, 55]. Эти результаты являются спорными из-за нескольких технических проблем, включая методы выборки и нормализации.Наш анализ открытых источников данных о пациентах с БП также подтвердил это. Следовательно, инфицирование CVB3 у пациентов может свидетельствовать о начале БП.
Когда мы инфицировали мышей CVB3, инфекция сначала появлялась в области нескольких анатомических структур вдоль желудочков, что указывает на периферический путь заражения CVB3 в ЦНС. Инфекция нейронов и микроглии со временем распространилась на другие регионы. В нейронах CVB3 наблюдали в гиппокампе, латеральном таламусе и среднем мозге мышей на 4-й день PI.Хотя мы наблюдали совместную локализацию CVB3 с α-syn in vitro , мы не смогли наблюдать совместную локализацию в головном мозге мышей, инфицированных CVB3. Вместо этого мы наблюдали накопление α-син в телах нейронов, расположенных в среднем мозге мышей WT на 28-й день PI, и больше нейронов, содержащих цитозольное накопление α-syn, наблюдали у мышей α-syn TG на 7-й день PI. Кроме того, вестерн-блот-анализ показал, что экспрессия pSer129 α-syn была немного увеличена в головном мозге, инфицированном CVB3.В предыдущем сообщении [41] авторы предположили, что одних триггеров обычно недостаточно для развития БП. Триггеры часто действуют преходяще, при этом триггерное событие длится несколько недель или месяцев и происходит относительно рано в жизни людей, у которых развивается БП. Соответственно, инфекция CVB3 может действовать транзиторно. Он сам по себе не может индуцировать образование LB в головном мозге, в отличие от наблюдений in vitro . В качестве альтернативы, агрегация нейронов α-син, наблюдаемая в нашей системе in vivo , может не достигать той же стадии созревания, что и LB PD, поскольку наша модельная система in vivo не подходит для длительных наблюдений из-за высокой смертности C57BL/ 6 мышей в ответ на инфекцию CVB3 [56].Долгосрочные последствия инфекции CVB3 в ЦНС в значительной степени неизвестны. Однако эти вирусы сохраняются на крайне низком уровне в ЦНС взрослых [57].
Кроме того, вестерн-блот-анализ показал, что экспрессия pSer129 α-syn была немного увеличена в головном мозге, инфицированном CVB3.В предыдущем сообщении [41] авторы предположили, что одних триггеров обычно недостаточно для развития БП. Триггеры часто действуют преходяще, при этом триггерное событие длится несколько недель или месяцев и происходит относительно рано в жизни людей, у которых развивается БП. Соответственно, инфекция CVB3 может действовать транзиторно. Он сам по себе не может индуцировать образование LB в головном мозге, в отличие от наблюдений in vitro . В качестве альтернативы, агрегация нейронов α-син, наблюдаемая в нашей системе in vivo , может не достигать той же стадии созревания, что и LB PD, поскольку наша модельная система in vivo не подходит для длительных наблюдений из-за высокой смертности C57BL/ 6 мышей в ответ на инфекцию CVB3 [56].Долгосрочные последствия инфекции CVB3 в ЦНС в значительной степени неизвестны. Однако эти вирусы сохраняются на крайне низком уровне в ЦНС взрослых [57]. Тем не менее, присутствие вирусной РНК само по себе является потенциально патогенным в некоторых случаях, таких как шизофрения [58] и боковой амиотрофический склероз [59]. Очень низкий титр вируса в ЦНС может быть недостаточным для наблюдения за распространением инфекции CVB3 и колокализацией между CVB3 и α-syn в головном мозге мышей, инфицированных CVB3, с помощью ИГХ.Интересно, что у мышей BALB/c, инфицированных CVB3, которые более восприимчивы к хронической инфекции CVB3, по сообщениям, наблюдалась агрегация TDP-43 в области гиппокампа через 90 дней PI [60]. Кроме того, сообщалось, что цитозольные агрегаты, а также растворимые олигомеры, которые не наблюдались у здоровых людей, наблюдались в сердце пациентов с дилатационной кардиомиопатией, которая, как предполагается, вызвана инфекциями CVB3 [61, 62]. Слитый островковый амилоидный полипептид конго красный также был обнаружен в биоптатах поджелудочной железы у пациентов с диабетом 1 типа, что позволяет предположить, что образование этих агрегатов может быть вызвано энтеровирусами [63].
Тем не менее, присутствие вирусной РНК само по себе является потенциально патогенным в некоторых случаях, таких как шизофрения [58] и боковой амиотрофический склероз [59]. Очень низкий титр вируса в ЦНС может быть недостаточным для наблюдения за распространением инфекции CVB3 и колокализацией между CVB3 и α-syn в головном мозге мышей, инфицированных CVB3, с помощью ИГХ.Интересно, что у мышей BALB/c, инфицированных CVB3, которые более восприимчивы к хронической инфекции CVB3, по сообщениям, наблюдалась агрегация TDP-43 в области гиппокампа через 90 дней PI [60]. Кроме того, сообщалось, что цитозольные агрегаты, а также растворимые олигомеры, которые не наблюдались у здоровых людей, наблюдались в сердце пациентов с дилатационной кардиомиопатией, которая, как предполагается, вызвана инфекциями CVB3 [61, 62]. Слитый островковый амилоидный полипептид конго красный также был обнаружен в биоптатах поджелудочной железы у пациентов с диабетом 1 типа, что позволяет предположить, что образование этих агрегатов может быть вызвано энтеровирусами [63]. Соответственно, индуцированное вирусом образование внутриклеточных белковых включений не ограничивается нейронами; скорее, это общее явление.
Соответственно, индуцированное вирусом образование внутриклеточных белковых включений не ограничивается нейронами; скорее, это общее явление.
В этом исследовании мы наблюдали, что α-син регулирует репликацию CVB3. Сверхэкспрессия α-syn приводила к усилению репликации CVB3 и индуцированной CVB3 цитотоксичности. Мы подтвердили это in vivo в мозге мышей α-syn TG. Репликация CVB3 была увеличена, а распределение CVB3-инфицированных областей также увеличилось у мышей α-syn TG по сравнению с таковым у контрольных мышей.Кроме того, инфекция CVB3 индуцировала гибель нейронов, включая потерю дофаминергических нейронов в черной субстанции. Интересно, что подтверждает наше наблюдение, что у пациентов с хроническим энцефалитом EV71, симптомы которых сохранялись более 2 месяцев, обнаруживалось поражение большей части среднего мозга, включая черную субстанцию [64]. Гибель дофаминергических нейронов черной субстанции также ускорялась у мышей α-syn TG. Кроме того, выживаемость мышей α-syn TG была меньше по сравнению с мышами дикого типа. Предыдущий отчет показал, что экспрессия α-syn ингибирует рост и репликацию ВЗН, что приводит к увеличению смертности мышей с нокаутом α-syn [21], что противоречит нашим выводам, которые демонстрируют, что мыши α-syn TG демонстрируют усиленную репликацию CVB3 и более низкая выживаемость после инфекции CVB3. Мы не можем полностью объяснить расхождение между этими результатами. Остаточное количество α-син в головном мозге может быть важным для регуляции вирусной инфекции. Тем не менее, это может быть связано с различными путями, связанными с аутофагией, задействованными вирусом.Предыдущие исследования показали, что CVB3 использует аутофагосомы в качестве своих центров репликации путем ингибирования слияния аутофагосом с лизосомами [28, 65, 66]. Индукция аутофагии в нейронах также была связана с усилением репликации CVB3 [65–67]. Увеличение экспрессии α-син ускоряет аутофагический поток. Также сообщалось, что сверхэкспрессия α-syn достаточна для нарушения созревания аутофагосом в модельной системе Drosophila [68].
Предыдущий отчет показал, что экспрессия α-syn ингибирует рост и репликацию ВЗН, что приводит к увеличению смертности мышей с нокаутом α-syn [21], что противоречит нашим выводам, которые демонстрируют, что мыши α-syn TG демонстрируют усиленную репликацию CVB3 и более низкая выживаемость после инфекции CVB3. Мы не можем полностью объяснить расхождение между этими результатами. Остаточное количество α-син в головном мозге может быть важным для регуляции вирусной инфекции. Тем не менее, это может быть связано с различными путями, связанными с аутофагией, задействованными вирусом.Предыдущие исследования показали, что CVB3 использует аутофагосомы в качестве своих центров репликации путем ингибирования слияния аутофагосом с лизосомами [28, 65, 66]. Индукция аутофагии в нейронах также была связана с усилением репликации CVB3 [65–67]. Увеличение экспрессии α-син ускоряет аутофагический поток. Также сообщалось, что сверхэкспрессия α-syn достаточна для нарушения созревания аутофагосом в модельной системе Drosophila [68]. Эта среда способствует репликации CVB3.Напротив, известно, что аутофагия ингибирует репликацию ВЗН [69, 70]. Соответственно, это может быть специфичным для вируса, и аутофагия у обоих вирусов может объяснить это несоответствие.
Эта среда способствует репликации CVB3.Напротив, известно, что аутофагия ингибирует репликацию ВЗН [69, 70]. Соответственно, это может быть специфичным для вируса, и аутофагия у обоих вирусов может объяснить это несоответствие.
Эпидемиологическая связь предполагает, что воздействие вируса с течением времени может увеличить риск БП, хотя неясно, вызывает ли БП какая-либо конкретная вирусная инфекция. Это может быть связано с прямой вирусной цитотоксичностью или вирусным воспалением [13, 71, 72]. Вирусные инфекции гриппа вызывают симптомы паркинсонизма и значительное увеличение фосфорилирования и агрегации α-син [73, 74].Повторная вирусная инфекция может индуцировать экспрессию α-син и/или агрегацию α-син, а хроническая вирусная инфекция вызывает дальнейшее воспаление, которое может инициировать и приводить к прогрессированию БП. Точно так же мы наблюдали, что α-syn реагировал на инфекцию CVB3. Инфекция CVB3 регулировала экспрессию и агрегацию α-syn. α-Syn может функционировать как защитный механизм против вирусной инфекции.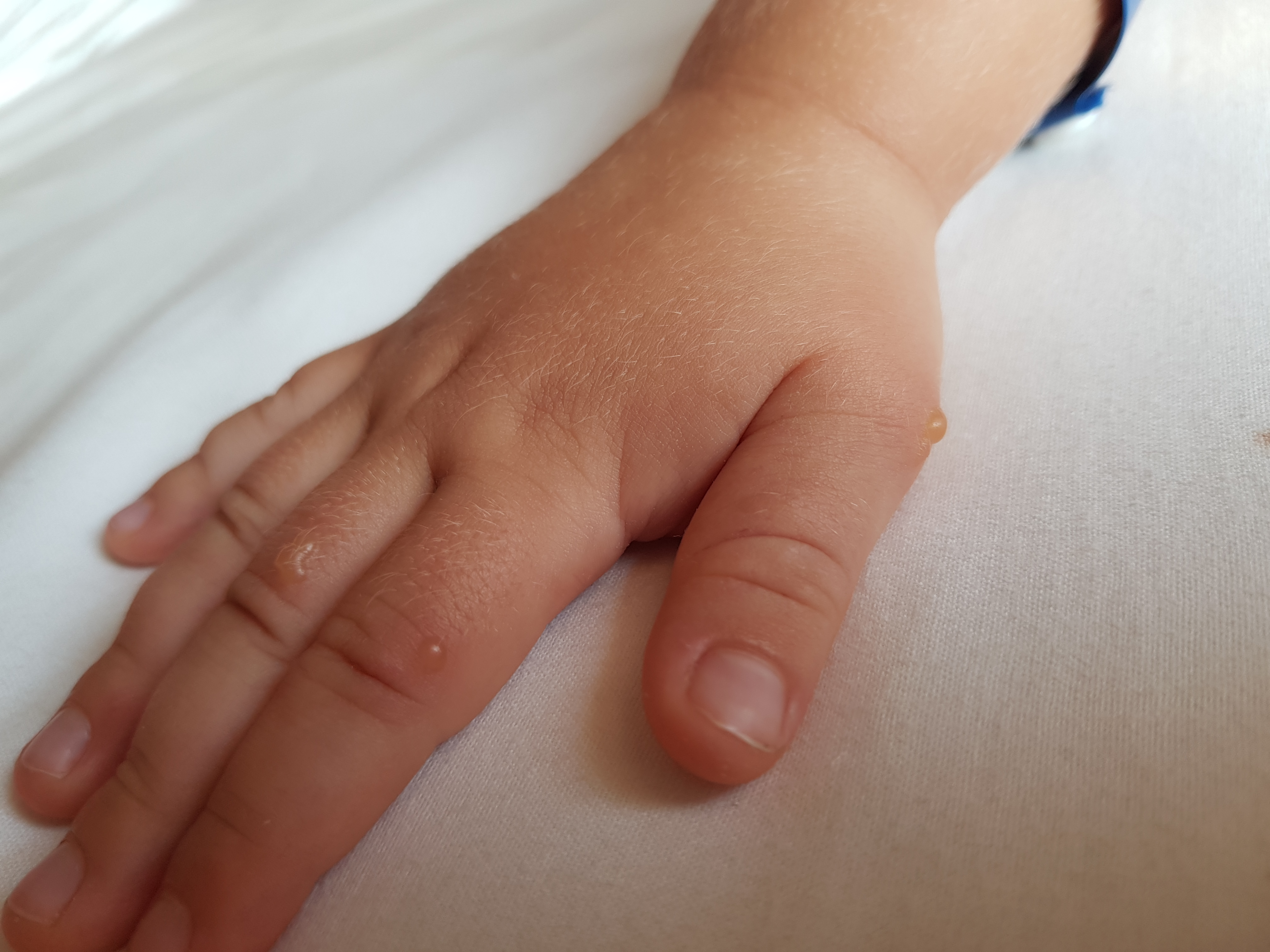 Тот факт, что инфекция CVB3 связана с α-сином, предполагает неожиданную роль α-сина в патогенезе БП. Необходимы дальнейшие исследования, чтобы изучить это более подробно.
Тот факт, что инфекция CVB3 связана с α-сином, предполагает неожиданную роль α-сина в патогенезе БП. Необходимы дальнейшие исследования, чтобы изучить это более подробно.
В заключение мы исследовали взаимосвязь между инфекцией CVB3 и α-syn. Инфекция CVB3 индуцирует образование включений, связанных с α-сином, в нейронах, которые могут действовать как триггер для БП. Эти тельца включения содержали сгруппированные органеллы, включая поврежденные митохондрии с фибриллами α-син. Сверхэкспрессия α-Syn ускоряет образование телец включения и индуцирует более концентрические тельца включения. Мозг мышей, инфицированных CVB3, содержал агрегаты α-син в теле клетки среднего мозга.Данные показывают, что инфекция CVB3 блокирует позднюю стадию аутофагии и индуцирует образование телец включения, содержащих фибриллы α-syn. Сверхэкспрессия α-syn способствует репликации CVB3 и связанной с ней цитотоксичности. Выживаемость мышей α-syn TG была низкой. Репликация CVB3 была более обширной у этих мышей, что дополнительно индуцировало гибель нейронов, включая потерю дофаминергических нейронов. Сверхэкспрессия α-Syn ускоряет поток аутофагии, что способствует репликации CVB3. В совокупности наши результаты проясняют механизм образования LB и патогенез БП, связанной с инфекцией CVB3.
Сверхэкспрессия α-Syn ускоряет поток аутофагии, что способствует репликации CVB3. В совокупности наши результаты проясняют механизм образования LB и патогенез БП, связанной с инфекцией CVB3.
Материалы и методы
Заявление об этике
Все процедуры проводились в соответствии с рекомендациями, установленными Комитетом по этике Медицинской школы Университета Аджу (IACUC № 2016–0047).
Антитела и реагенты
Антитела против α-syn были приобретены у Abcam (№ ab138501, Кембридж, Великобритания), BD Biosciences (№ 610786, Франклин Лейкс, Нью-Джерси) и Genetex (№ GTX112799, Санта-Барбара, Калифорния). Антитела против pSer129 α-syn (#ab51253), Tuj-1 (#ab18207) и энтеровируса 71 (#ab36367) были получены от Abcam.Антитело против VP1 CVB3 было приобретено у Millipore (#MAB948, Danvers, MA). Антитело против LC3 было приобретено у Sigma-Aldrich (№ L8918, Сент-Луис, Миссури). Антитело против p62 было приобретено у BD Biosciences (#610832). Антитело против расщепленной каспазы-3 было приобретено у Cell Signaling Technology (#9664, Беверли, Массачусетс). Антитело против Iba-1 было приобретено у Wako (№ 019–19741, Ричмонд, Вирджиния). Антитело против глиального фибриллярного кислого белка (GFAP) было приобретено у Neuromics (#RA22101, Montreal, QC).Антитело против глицеральдегид-3-фосфатдегидрогеназы (GAPDH) было приобретено у Santa Cruz Biotechnology (#SC-32233, Santa Cruz, CA). LysoTracker (№ L12492) и Lipofectamine 2000 (№ 11668–019) были получены от Invitrogen (Карлсбад, Калифорния). Ретиноевая кислота (RA, #R2625), бафиломицин A1 (Baf A1, #B1793), полиинозин: полицитидиловая кислота (poly IC, #P0913) и краситель Evans Blue Dye (EBD, #E2129) были приобретены у Sigma-Aldrich. Судан Блэк Б был приобретен в компании Tokyo Chemical Industry (№ 4197–2505, Токио, Япония).
Антитело против Iba-1 было приобретено у Wako (№ 019–19741, Ричмонд, Вирджиния). Антитело против глиального фибриллярного кислого белка (GFAP) было приобретено у Neuromics (#RA22101, Montreal, QC).Антитело против глицеральдегид-3-фосфатдегидрогеназы (GAPDH) было приобретено у Santa Cruz Biotechnology (#SC-32233, Santa Cruz, CA). LysoTracker (№ L12492) и Lipofectamine 2000 (№ 11668–019) были получены от Invitrogen (Карлсбад, Калифорния). Ретиноевая кислота (RA, #R2625), бафиломицин A1 (Baf A1, #B1793), полиинозин: полицитидиловая кислота (poly IC, #P0913) и краситель Evans Blue Dye (EBD, #E2129) были приобретены у Sigma-Aldrich. Судан Блэк Б был приобретен в компании Tokyo Chemical Industry (№ 4197–2505, Токио, Япония).
Животные
α-Syn трансгенных (TG) мышей со сверхэкспрессией α-syn человека под контролем промотора нейрон-специфической енолазы (NSE) (C57BL/6N-Tg (NSE-h a Syn) Korl) были предоставлены Национальным институтом пищевых продуктов и Оценка безопасности лекарственных средств (NIFDS, Чхонджу, Корея). В качестве контроля использовали однопометных мышей дикого типа (WT) или мышей WT C57BL/6N (DBL, Eumseong, Korea).
В качестве контроля использовали однопометных мышей дикого типа (WT) или мышей WT C57BL/6N (DBL, Eumseong, Korea).
Культура клеток
клетки α-Syn, mCherry и α-syn-mCherry со сверхэкспрессией SH-SY5Y были созданы, как описано ранее [75].Клетки α-Syn KO SH-SY5Y получали с использованием системы lentiCRISPR [76] (отожженные олигонуклеотиды α-syn человека; олиго 1: 5′-TGTAGGCTCCAAAAC-CAAGG-3′, олиго 2: 5′-CCTTGGTTTTGGAGCCTACA-3′). Олигонуклеотиды были разработаны с использованием онлайн-инструмента проектирования гРНК (https://chopchop.cbu.uib.no). Отдельные клоны были разведены из трансфицированной популяции, выделены и отобраны с использованием геномного секвенирования и вестерн-блоттинга. Клетки выращивали в среде Игла, модифицированной Дульбекко (DMEM) с добавлением 10% эмбриональной телячьей сыворотки (FBS) при 37°C во влажной атмосфере, содержащей 5% CO 2 и 95% воздуха.Первичные корковые нейроны выделяли у однодневных детенышей мышей WT и α-syn TG и культивировали в нейробазальной среде (№ 21103–049, Invitrogen) с GlutaMAX-I (№ 35050061, Thermo Fisher Scientific, Уолтем, Массачусетс). и добавку B-27 (№ 17504–044, Invitrogen) в течение 2 недель на покрытых поли-D-лизином (№ P7280, Sigma-Aldrich) предметных стеклах или чашках для культивирования клеток. Для дифференцировки клетки SH-SY5Y обрабатывали 50 мкМ RA в течение 5 дней.
и добавку B-27 (№ 17504–044, Invitrogen) в течение 2 недель на покрытых поли-D-лизином (№ P7280, Sigma-Aldrich) предметных стеклах или чашках для культивирования клеток. Для дифференцировки клетки SH-SY5Y обрабатывали 50 мкМ RA в течение 5 дней.
Инфекция вирусом Коксаки В3 (CVB3)
Вариант h4 CVB3, штамм Вудраффа и EGFP-CVB3 были любезным подарком от Dr.Э. Чжон (Медицинский центр Самсунг, Сеул, Корея). Вирус размножали в клетках HeLa. Вирусные титры определяли с помощью анализа бляшек [77]. Клетки SH-SY5Y инкубировали с CVB3 при множественности заражения 1 (MOI) в среде DMEM без сыворотки в течение 1 часа, а затем инкубировали в среде DMEM с добавлением 10% FBS в течение 30 часов. Для первичных нейронов клетки инкубировали с указанным MOI CVB3 в среде для культивирования нейронов в течение 24 часов. В модели in vivo самцов мышей в возрасте от 8 до 11 недель заражали внутрибрюшинной (IP) инъекцией 1.0 × 10 6 бляшкообразующих единиц (БОЕ) CVB3 в 100 мкл фосфатно-солевого буфера (PBS).
Подготовка ткани
Мышей анестезировали и транскардиально перфузировали сначала перфузионным раствором, содержащим 0,5% нитрата натрия и 10 ЕД/мл гепарина, а затем 4% параформальдегидом в 0,1 М фосфатном буфере (ФБ; рН 7,2). Мозги вначале хранили в 4% параформальдегиде в течение 24 ч при 4°С, а затем в 30% растворе сахарозы до их оседания. Для полимеразной цепной реакции с обратной транскрипцией (RT-PCR) и вестерн-блоттинга мышей перфузировали перфузионным раствором в течение 2 минут, и каждый орган хранили при -70 ° C до использования.Для иммуноокрашивания шесть отдельных серий корональных срезов головного мозга размером 30 мкм делали с помощью криостата (модель CM3050S, Leica, Wetzlar, Германия) и хранили в маточном растворе антифриза (PB, содержащем 30% глицерина и 30% этиленгликоля, pH 7,2). ) при 4°C перед экспериментами.
Иммуноцитохимия
Клетки, культивированные на покровных стеклах, трижды промывали PBS и фиксировали 4% параформальдегидом в течение 10 мин при комнатной температуре.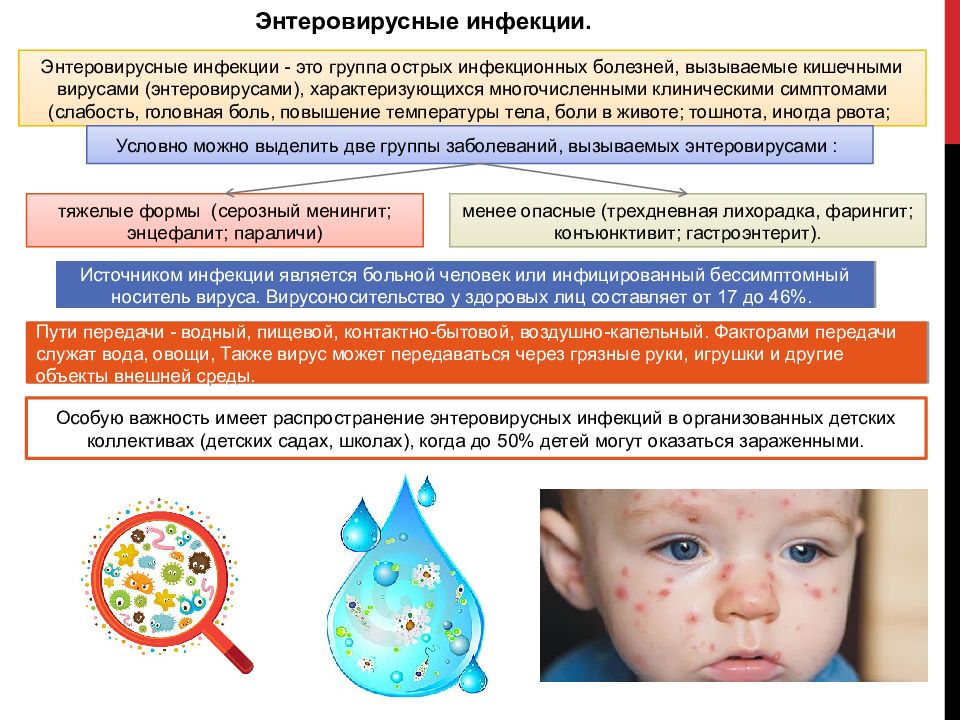 Фиксированные клетки несколько раз промывали PBS и пермеабилизировали PBS, содержащим 0.1% Triton X-100 в течение 5 мин при комнатной температуре. После промывки PBS клетки блокировали PBS, содержащим 1% бычьего сывороточного альбумина (БСА), в течение 1 ч при комнатной температуре, а затем инкубировали в течение ночи с указанными антителами при 4°C. Затем образцы инкубировали с конъюгированными вторичными антителами Alexa Fluor 488- (#A21202, #A21206) или Alexa Fluor 568- (#A10037, #A10042) (все от Invitrogen) в течение 1 часа, а затем окрашивали Hoechst в течение 5 минут. . Срезы монтировали и исследовали с помощью конфокальной микроскопии (LSM710, Carl Zeiss, Йена, Германия) в Центре трехмерной визуализации иммунной системы Университета Аджу.Живые клетки инкубировали с 50 нМ LysoTracker, приготовленным в DMEM с добавлением 10% FBS, в течение 30 мин. После окрашивания ядер Hoechst в течение 5 минут живые клетки наблюдали с помощью конфокальной микроскопии.
Фиксированные клетки несколько раз промывали PBS и пермеабилизировали PBS, содержащим 0.1% Triton X-100 в течение 5 мин при комнатной температуре. После промывки PBS клетки блокировали PBS, содержащим 1% бычьего сывороточного альбумина (БСА), в течение 1 ч при комнатной температуре, а затем инкубировали в течение ночи с указанными антителами при 4°C. Затем образцы инкубировали с конъюгированными вторичными антителами Alexa Fluor 488- (#A21202, #A21206) или Alexa Fluor 568- (#A10037, #A10042) (все от Invitrogen) в течение 1 часа, а затем окрашивали Hoechst в течение 5 минут. . Срезы монтировали и исследовали с помощью конфокальной микроскопии (LSM710, Carl Zeiss, Йена, Германия) в Центре трехмерной визуализации иммунной системы Университета Аджу.Живые клетки инкубировали с 50 нМ LysoTracker, приготовленным в DMEM с добавлением 10% FBS, в течение 30 мин. После окрашивания ядер Hoechst в течение 5 минут живые клетки наблюдали с помощью конфокальной микроскопии.
Вестерн-блоттинг
Образцы лизировали с использованием охлажденного льдом буфера RIPA (50 мМ Tris-HCl, pH 7,4, 0,5 % дезоксихолата натрия, 150 мМ NaCl, 0,1 % SDS, 1 % Triton X-100), содержащего смесь ингибиторов протеаз (№ 535140, Calbiochem , Дармштадт, Германия) и смесь ингибиторов фосфатазы (#P3200-001, GenDEPOT, Baker, TX). После кратковременной обработки ультразвуком лизаты центрифугировали при 16 000 g в течение 30 мин при 4°C и собирали супернатанты. В случае тканей мышей (мозг и сердце) после лизиса и гомогенизации образцов с помощью TRIzol (#TR118, Molecular Research Center Inc, Цинциннати, Огайо, США) белки выделяли в соответствии с протоколом производителя. Концентрации белка определяли с использованием пакета реагентов для анализа белков DC (№ 5000116, Bio-Rad, Hercules, CA). Белки разделяли с помощью SDS-PAGE, переносили на мембрану PVDF или NC и подвергали иммуноблотингу с указанными первичными антителами, а затем с конъюгированными с пероксидазой хрена вторичными антителами (#G-21040, Invitrogen или #111-035-003, Jackson ImmunoResearch, West Гроув, Пенсильвания).Затем белки визуализировали с использованием системы усиленной хемилюминесценции (ECL) (#LF-QC0101, AbFrontier, Сеул, Корея). Интенсивность полос определяли с помощью ImageJ (NIH, Bethesda, MD).
После кратковременной обработки ультразвуком лизаты центрифугировали при 16 000 g в течение 30 мин при 4°C и собирали супернатанты. В случае тканей мышей (мозг и сердце) после лизиса и гомогенизации образцов с помощью TRIzol (#TR118, Molecular Research Center Inc, Цинциннати, Огайо, США) белки выделяли в соответствии с протоколом производителя. Концентрации белка определяли с использованием пакета реагентов для анализа белков DC (№ 5000116, Bio-Rad, Hercules, CA). Белки разделяли с помощью SDS-PAGE, переносили на мембрану PVDF или NC и подвергали иммуноблотингу с указанными первичными антителами, а затем с конъюгированными с пероксидазой хрена вторичными антителами (#G-21040, Invitrogen или #111-035-003, Jackson ImmunoResearch, West Гроув, Пенсильвания).Затем белки визуализировали с использованием системы усиленной хемилюминесценции (ECL) (#LF-QC0101, AbFrontier, Сеул, Корея). Интенсивность полос определяли с помощью ImageJ (NIH, Bethesda, MD).
ОТ-ПЦР
Тотальную РНК выделяли из образцов с использованием TRIzol (#TR118, Molecular Research Center Inc. ) в соответствии с протоколом производителя. Тотальную РНК подвергали обратной транскрипции с использованием обратной транскриптазы AMV (#M0277L, New England Biolabs, Ipswich, MA). Уровни транскриптов генов-мишеней количественно определяли с использованием 2 X KAPA SYBR Fast Master Mix (#kk4602, Kapa Biosystems, Кейптаун, Южная Африка) с использованием системы ПЦР в реальном времени StepOnePlus (Applied Biosystems, Фостер-Сити, Калифорния).Для каждого гена-мишени уровень транскриптов нормализовали к уровню GAPDH и рассчитывали с использованием стандартного метода ΔΔCT. Полный список последовательностей праймеров представлен в дополнительной таблице S1.
) в соответствии с протоколом производителя. Тотальную РНК подвергали обратной транскрипции с использованием обратной транскриптазы AMV (#M0277L, New England Biolabs, Ipswich, MA). Уровни транскриптов генов-мишеней количественно определяли с использованием 2 X KAPA SYBR Fast Master Mix (#kk4602, Kapa Biosystems, Кейптаун, Южная Африка) с использованием системы ПЦР в реальном времени StepOnePlus (Applied Biosystems, Фостер-Сити, Калифорния).Для каждого гена-мишени уровень транскриптов нормализовали к уровню GAPDH и рассчитывали с использованием стандартного метода ΔΔCT. Полный список последовательностей праймеров представлен в дополнительной таблице S1.
Трансмиссионная электронная микроскопия (ПЭМ)
Контрольные образцы и инфицированные CVB3 образцы фиксировали 0,1 М буфером какодилата натрия (pH 7,4), содержащим 1% формальдегида / 2% глутаральдегида, в течение 30 мин при 4°C [78]. Образцы дважды промывали холодным PBS, затем фиксировали смесью 1% четырехокиси осмия и 1% феррицианида калия, обезвоживали в спирте определенной степени и заливали в смолу Durcupan ACM (Fluka, Yongin, Корея). Смолой были получены ультратонкие срезы, помещенные на медные сетки и контрастно окрашенные уранилацетатом и цитратом свинца. Образцы наблюдали с помощью просвечивающего электронного микроскопа Sigma 500 (Carl Zeiss, Йена, Германия) в Центре трехмерной визуализации иммунной системы Университета Аджу.
Смолой были получены ультратонкие срезы, помещенные на медные сетки и контрастно окрашенные уранилацетатом и цитратом свинца. Образцы наблюдали с помощью просвечивающего электронного микроскопа Sigma 500 (Carl Zeiss, Йена, Германия) в Центре трехмерной визуализации иммунной системы Университета Аджу.
Анализ морфологии митохондрий
Морфологию митохондрий в ПЭМ-изображениях анализировали, как описано ранее [33]. По типу митохондриальной перестройки выделены четыре категории (тип I-IV).Тип I состоял из митохондриальных крист, которые были правильными, плотно упакованными и продольно ориентированными. Тип II включает митохондрии аномальной формы или неоднородного размера, кристы неправильной формы, лишенные ориентации и плотности. Тип III включает митохондрии различной формы и размеров с прерывистой наружной мембраной, фрагментированными кристами и набухшим матриксом. Тип IV включал митохондрии с разорванной наружной мембраной, без крист и миелиноподобной трансформацией..jpg)
Анализ цитотоксичности
Анализы цитотоксичностивыполняли с использованием набора для анализа цитотоксичности ЛДГ (№ K311-400, BioVision; Маунтин-Вью, Калифорния) в соответствии с протоколом производителя.После инфицирования CVB3 в течение 30 ч 50 мкл среды и раствора набора для анализа смешивали в лунках оптически прозрачного 96-луночного планшета. Поглощение измеряли в течение 10 минут при 490 нм с помощью считывателя ELISA (Molecular Device, Wokingham, UK)
Проточная цитометрия
Проточную цитометрию проводили, как описано ранее [79]. После заражения EGFP-CVB3 в указанные моменты времени клетки выделяли и фиксировали ресуспендированием в 4% параформальдегиде в PBS в течение ночи. После промывания PBS и ресуспендирования в 1% BSA в PBS клетки анализировали с использованием сортировщика клеток FACS Aria III (BD, Franklin Lakes, NJ) в Центре трехмерной визуализации иммунной системы Университета Аджу.Данные были проанализированы с использованием программного обеспечения Flowing версии 2. 5.1 (Turku Bioscience, Турку, Финляндия).
5.1 (Turku Bioscience, Турку, Финляндия).
Иммуногистохимия (ИГХ)
Каждый серийный срез в каждом наборе собирали и промывали PBS, содержащим 0,2% Triton X-100 (PBST). После блокировки 1% BSA в PBST срезы инкубировали в течение ночи при комнатной температуре с указанными первичными антителами. Образцы инкубировали с конъюгированными вторичными антителами Alexa Fluor 488- (#A21202, #A21206) или Alexa Fluor 568- (#A10037, #A10042) (все от Invitrogen) в течение 1 часа, а затем окрашивали с помощью Hoechst в течение 10 минут.После установки на предметные стекла срезы обрабатывали раствором судана черного Б (0,1% в 70% этаноле) для ингибирования автофлуоресценции тканей мыши. Изображения были получены с помощью конфокального микроскопа (LSM710, Carl Zeiss, Йена, Германия) или флуоресцентного микроскопа (Axioscan Z1, Carl Zeiss, Йена, Германия) в Центре трехмерной визуализации иммунной системы Университета Аджу.
Анализ плотности клеток
Полушария мышей использовали для обнаружения VP1 методом ОТ-ПЦР. Противоположные полушария использовались для анализа плотности клеток. Были проанализированы полушария трех мышей, у которых уровень VP1 в мозге был близок к среднему. Мы измерили количество клеток (нейронов) и плотность клеток (микроглии) на конкретную анатомическую структуру. VP1-положительные клетки, объединенные Tuj-1, подсчитывали из трех эквивалентных мест для нейронов, а Iba-1, объединенные VP1-положительные клетки, из шести эквивалентных мест для микроглии вдоль рострокаудальной оси. Клетки были отмечены предложенным цветом.Микроглию подсчитывали с использованием модуля роста нейритов MetaMorph (Molecular Devices). Плотность получали путем деления подсчитанного значения на площадь каждой анатомической структуры. Анатомическая структура была разделена на кору головного мозга, ядра головного мозга, фимбрии, внутреннюю капсулу, таламус, гипоталамус и средний мозг. В обонятельной области кора головного мозга разделялась на изокортекс и обонятельную области, а гиппокамп отделялся от коры головного мозга в отделе, включающем гиппокамп.
Противоположные полушария использовались для анализа плотности клеток. Были проанализированы полушария трех мышей, у которых уровень VP1 в мозге был близок к среднему. Мы измерили количество клеток (нейронов) и плотность клеток (микроглии) на конкретную анатомическую структуру. VP1-положительные клетки, объединенные Tuj-1, подсчитывали из трех эквивалентных мест для нейронов, а Iba-1, объединенные VP1-положительные клетки, из шести эквивалентных мест для микроглии вдоль рострокаудальной оси. Клетки были отмечены предложенным цветом.Микроглию подсчитывали с использованием модуля роста нейритов MetaMorph (Molecular Devices). Плотность получали путем деления подсчитанного значения на площадь каждой анатомической структуры. Анатомическая структура была разделена на кору головного мозга, ядра головного мозга, фимбрии, внутреннюю капсулу, таламус, гипоталамус и средний мозг. В обонятельной области кора головного мозга разделялась на изокортекс и обонятельную области, а гиппокамп отделялся от коры головного мозга в отделе, включающем гиппокамп. В случае ядер головного мозга полосатое тело и паллидум были в значительной степени разделены, а полосатое тело снова было разделено на дорсальную область, комплекс латеральных перегородок и вентральную область. Количество ТГ-позитивных клеток в полушариях головного мозга контрольных и инфицированных CVB3 мышей подсчитывали вручную на иммуногистохимических изображениях (Axioscan Z1, Carl Zeiss, Jena, Germary). Среднее количество срезов SNpc для мыши составляло от 5 до 6 в серии 1: 6 в наших экспериментальных условиях (срезы 30 мкм). Цифры представляют собой общее количество TH-положительных клеток на мышь, умноженное на 6 для расчета оценки популяции.
В случае ядер головного мозга полосатое тело и паллидум были в значительной степени разделены, а полосатое тело снова было разделено на дорсальную область, комплекс латеральных перегородок и вентральную область. Количество ТГ-позитивных клеток в полушариях головного мозга контрольных и инфицированных CVB3 мышей подсчитывали вручную на иммуногистохимических изображениях (Axioscan Z1, Carl Zeiss, Jena, Germary). Среднее количество срезов SNpc для мыши составляло от 5 до 6 в серии 1: 6 в наших экспериментальных условиях (срезы 30 мкм). Цифры представляют собой общее количество TH-положительных клеток на мышь, умноженное на 6 для расчета оценки популяции.
Статистический анализ
Все значения экспериментальных данных выражены как среднее значение ± стандартное отклонение. Статистическую значимость оценивали с использованием непарного t-критерия, однофакторного дисперсионного анализа или двухфакторного дисперсионного анализа с использованием программного обеспечения Graphpad Prism (GraphPad, La Jolla, CA). Линейная регрессия была выполнена для анализа корреляции между mCherry и сигналом EGFP с использованием программного обеспечения GraphPad Prism.
Линейная регрессия была выполнена для анализа корреляции между mCherry и сигналом EGFP с использованием программного обеспечения GraphPad Prism.
Вспомогательная информация
S1 Рис. Окрашивание H&E и анализ цитотоксичности между контрольными и CVB3-зараженными клетками dSH-SY5Y и исследование экспрессии α-syn между контрольными и CVB3-зараженными образцами, старым контролем и пациентами с БП.
(A) Иммуноцитохимические (ICC) изображения клеток α-syn KO dSH-SY5Y, инфицированных CVB3 (MOI 0,25) (красный) в течение 24 часов. Масштабная линейка указывает 10 мкм. (B) Окрашивание H&E контрольных клеток α-syn OE dSH-SY5Y, инфицированных CVB3 (MOI 0,25) в течение 24 часов. Масштабная линейка указывает 20 мкм. Стрелки указывают на эозин-положительные включения клеток, инфицированных CVB3. (C) Анализ цитотоксичности клеток WT и α-syn OE dSH-SY5Y, инфицированных 0,25 MOI CVB3 в течение 24 часов, с помощью анализа LDH. Значения получены из трех независимых экспериментов (n = 3), одностороннего теста ANOVA с тестом множественных сравнений Тьюки.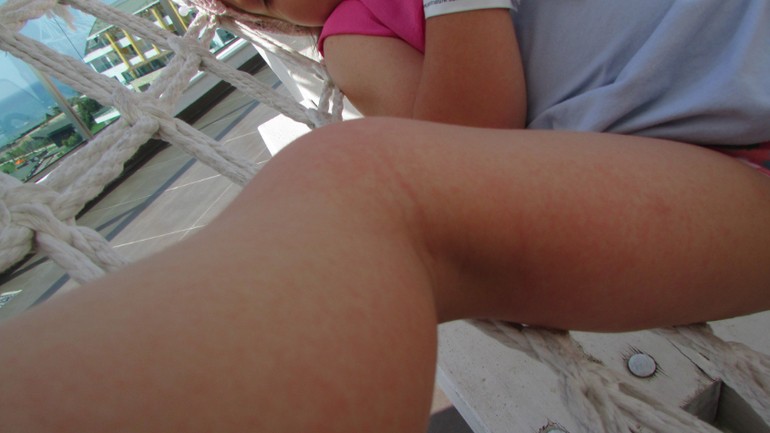 (D) Относительные уровни экспрессии нескольких мРНК в контрольных и инфицированных CVB3 клетках dSH-SY5Y (MOI 0,25) в течение 24 ч [24–26]. Значения получены из трех независимых экспериментов (n = 3), непарного t-критерия. (E) Комплексный анализ экспрессии генов (GEO) относительных уровней α-син в контрольных (n = 3) и CVB3-инфицированных (внутрибрюшинная (IP) инъекция 400 БОЕ) мышах (n = 3) сердцах на 4-й день после заражения (PI) у 4 типов линий мышей с помощью профилирования экспрессии массива (GSE19496). Дифференциальную экспрессию генов (DGE) анализировали с помощью пакета Limma в R.Значение P для каждого анализа было следующим: штамм A/J (9,20E -07 ), штамм B10.A-h3a (7,54E -06 ), штамм B6.Chr 3A/J (3,47E — 05 ), штамм CSS3 (6.66E -09 ). (F) Относительные уровни α-син в посмертном мозге нормальных (n = 9) и пациентов с БП (n = 16), исследованных с помощью профилирования экспрессии массива (GSE7621). ДЭГ анализировали с помощью пакета limma в R.
(D) Относительные уровни экспрессии нескольких мРНК в контрольных и инфицированных CVB3 клетках dSH-SY5Y (MOI 0,25) в течение 24 ч [24–26]. Значения получены из трех независимых экспериментов (n = 3), непарного t-критерия. (E) Комплексный анализ экспрессии генов (GEO) относительных уровней α-син в контрольных (n = 3) и CVB3-инфицированных (внутрибрюшинная (IP) инъекция 400 БОЕ) мышах (n = 3) сердцах на 4-й день после заражения (PI) у 4 типов линий мышей с помощью профилирования экспрессии массива (GSE19496). Дифференциальную экспрессию генов (DGE) анализировали с помощью пакета Limma в R.Значение P для каждого анализа было следующим: штамм A/J (9,20E -07 ), штамм B10.A-h3a (7,54E -06 ), штамм B6.Chr 3A/J (3,47E — 05 ), штамм CSS3 (6.66E -09 ). (F) Относительные уровни α-син в посмертном мозге нормальных (n = 9) и пациентов с БП (n = 16), исследованных с помощью профилирования экспрессии массива (GSE7621). ДЭГ анализировали с помощью пакета limma в R. Значение P = 0,002
Значение P = 0,002
https://doi.org/10.1371/journal.ppat.1010018.s002
(TIF)
S2 Рис.Инфекция энтеровирусом 71 не образует аутофагосомы, колокализованные с α-син.
ICC-изображения клеток WT dSH-SY5Y, неинфицированных (контроль) или инфицированных энтеровирусом 71 (EV71) (MOI 1) за 24 часа. Клетки иммуноокрашивали указанными антителами. Белые стрелки указывают на LC3-положительные агрегаты.
https://doi.org/10.1371/journal.ppat.1010018.s003
(TIF)
S3 Рис. Повышенный уровень α-син увеличивает поток аутофагии.
(A) Графики вулканов DEG между контролем и средой с повышенным содержанием α-син.(Наборы данных GEO, соответствующие сверхэкспрессии α-syn человека с использованием лентивирусного вектора в нейронах среднего мозга мыши (GSE70368), индуцированных плюрипотентных стволовых клетках (iPSC) тройного семейства α-syn (SNCA) (GSE30792) и ткани стриатума мыши были проанализированы человеческие α-син трансгенные (TG) мыши (GSE116010). Светло- и темно-серые точки обозначают гены, экспрессия которых изменялась незначительно и достоверно соответственно. Красные точки представляют гены с повышенной регуляцией, а синие точки представляют гены с отрицательной регуляцией.GSE70368 и GSE116010 анализировали с использованием пакета Deseq2, а GSE30792 анализировали с использованием пакета Limma в R. (B) График GO для «положительной регуляции макроаутофагии» (GO:0016239), «созревания аутофагосом» (GO:0097352) и «лизосомы». организация» (GO:0007040) в анализе генной онтологии (GO) GSE70368, GSE30792 и GSE116010, которые были выше, чем контрольные в каждом наборе.
Светло- и темно-серые точки обозначают гены, экспрессия которых изменялась незначительно и достоверно соответственно. Красные точки представляют гены с повышенной регуляцией, а синие точки представляют гены с отрицательной регуляцией.GSE70368 и GSE116010 анализировали с использованием пакета Deseq2, а GSE30792 анализировали с использованием пакета Limma в R. (B) График GO для «положительной регуляции макроаутофагии» (GO:0016239), «созревания аутофагосом» (GO:0097352) и «лизосомы». организация» (GO:0007040) в анализе генной онтологии (GO) GSE70368, GSE30792 и GSE116010, которые были выше, чем контрольные в каждом наборе.
https://doi.org/10.1371/journal.ppat.1010018.s004
(TIF)
S4 Рис. Экспрессия α-Syn в мозге мышей WT и TG, CVB3-инфицированных нейронах и микроглии.
(A) Вестерн-блоттинг проводили с использованием лизатов мозга контрольных и α-syn TG мышей. Уровни белка определяли количественно с помощью денситометрии. *** P <0,001, непарный t-критерий. ИГХ-изображения мозга мышей, инфицированных CVB3, на 4, 7 и 28 день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Показаны Tuj-1 (таламус) (B) и Iba-1 (обонятельная луковица) (C) положительные клетки, которые были локализованы вместе с VP1 у инфицированных CVB3 мышей WT и α-syn TG. Белые стрелки указывают на CVB3-инфицированные Tuj-1 положительные клетки. Синий указывает на DAPI.Масштабная линейка указывает 20 мкм.
ИГХ-изображения мозга мышей, инфицированных CVB3, на 4, 7 и 28 день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). Показаны Tuj-1 (таламус) (B) и Iba-1 (обонятельная луковица) (C) положительные клетки, которые были локализованы вместе с VP1 у инфицированных CVB3 мышей WT и α-syn TG. Белые стрелки указывают на CVB3-инфицированные Tuj-1 положительные клетки. Синий указывает на DAPI.Масштабная линейка указывает 20 мкм.
https://doi.org/10.1371/journal.ppat.1010018.s005
(TIF)
S5 Рис. Активация микроглии инфекцией CVB3 в головном мозге мышей.
(A) ICC-изображения мозга контрольных и CVB3-инфицированных мышей на 7-й день PI (в/б инъекция 1,0 × 10 6 БОЕ CVB3) и увеличенные изображения со схематической диаграммой микроглии для анализа (черная линия). Масштабная линейка указывает 10 мкм. (B) Морфологический анализ VP1-положительной микроглии в головном мозге CVB3-инфицированных мышей на 7-й день PI.Относительные уровни размера клеточного тела, длины отростка и отношения размаха между контролем и колокализованной VP1 микроглией. Значения получены из микроглии, случайно выбранной из указанной анатомической структуры трех мышей. * P <0,05, *** P <0,001, непарный t-критерий.
Значения получены из микроглии, случайно выбранной из указанной анатомической структуры трех мышей. * P <0,05, *** P <0,001, непарный t-критерий.
https://doi.org/10.1371/journal.ppat.1010018.s006
(TIF)
S6 Рис. Цитотоксичность, индуцированная CVB3, в мозге мышей.
ICC-изображения головного мозга мышей контрольной группы и мышей, инфицированных CVB3, на 28-й день PI (интраперитонеальная инъекция 1.0 × 10 6 БОЕ CVB3). Белые стрелки указывают на расщепленные каспаза-3 (с-каспаза-3) положительные клетки. Масштабная линейка указывает 10 мкм. Синий указывает на DAPI.
https://doi.org/10.1371/journal.ppat.1010018.s007
(TIF)
S7 Рис. Повреждение тканей и репликация CVB3 в органах мышей WT и α-syn TG.
(A) ИГХ-изображения окрашивания синего Эванса и анализа интенсивности контрольных (n = 3), CVB3-инфицированных мышей WT (n = 4) и α-syn TG мышей (n = 6) сердца на 7-й день PI (IP инъекция 1.0 × 10 6 БОЕ CVB3). Масштабная линейка указывает 500 мкм. Односторонний тест ANOVA с тестом множественных сравнений Тьюки. (B) Относительные уровни VP1 между сердцами мышей дикого типа (n = 4) и α-syn TG (n = 7) на 7-й день PI (внутрибрюшинная инъекция 1,0 × 10 6 БОЕ CVB3). Был проведен непарный t-тест. (C) Окрашивание сердца, поджелудочной железы, печени и селезенки гематоксилином и эозином у контрольных и CVB3-инфицированных мышей WT и α-syn TG на 7-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). (D) Вестерн-блоттинг проводили с использованием лизатов головного мозга, сердца и поджелудочной железы контрольных мышей и мышей α-syn TG.
Масштабная линейка указывает 500 мкм. Односторонний тест ANOVA с тестом множественных сравнений Тьюки. (B) Относительные уровни VP1 между сердцами мышей дикого типа (n = 4) и α-syn TG (n = 7) на 7-й день PI (внутрибрюшинная инъекция 1,0 × 10 6 БОЕ CVB3). Был проведен непарный t-тест. (C) Окрашивание сердца, поджелудочной железы, печени и селезенки гематоксилином и эозином у контрольных и CVB3-инфицированных мышей WT и α-syn TG на 7-й день PI (интраперитонеальная инъекция 1,0 × 10 6 БОЕ CVB3). (D) Вестерн-блоттинг проводили с использованием лизатов головного мозга, сердца и поджелудочной железы контрольных мышей и мышей α-syn TG.
https://doi.org/10.1371/journal.ppat.1010018.s008
(TIF)
Благодарности
Мы благодарим доктора Э. Чона (Медицинский центр Samsung, Сеул, Корея) за любезно предоставленный вариант h4 CVB3, штамм Вудраффа и EGFP-CVB3. Мы благодарим профессора Сан-Янг Чанг (Фармацевтический колледж Университета Аджу, Сувон, Корея) за любезно предоставленный EV71. Мы также благодарим NIFDS за предоставление мышей C56BL/6-Tg (NSE-haSyn)Korl и сопутствующую информацию.
Мы также благодарим NIFDS за предоставление мышей C56BL/6-Tg (NSE-haSyn)Korl и сопутствующую информацию.
Каталожные номера
- 1.Poewe W, Seppi K, Tanner CM, Halliday GM, Brundin P, Volkmann J, et al. Болезнь Паркинсона. Обзоры природы. Буквы о болезнях. 2017;3:17013. пмид:28332488
- 2. Калия Л.В., Ланг А.Е. Болезнь Паркинсона. Ланцет. 2015;386(9996):896–912. Эпб 2015/04/24. пмид: 251.
- 3. Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. Альфа-синуклеин в тельцах Леви. Природа. 1997;388(6645):839–40. Эпб 1997/08/28. пмид:9278044.
- 4.Эшбах Дж., Данцер К.М. α-Синуклеин при болезни Паркинсона: патогенная функция и перевод на модели животных. Нейродегенеративные заболевания. 2014;14(1):1–17. пмид:24080741
- 5. Маллин С., Шапира А. Генетика болезни Паркинсона. Британский медицинский бюллетень. 2015;114(1):39–52. пмид:25995343
- 6.
Наллс М.А., Панкрац Н. , Лилль К.М., До К.Б., Эрнандес Д.Г., Саад М. и соавт. Крупномасштабный метаанализ данных об ассоциации всего генома выявил шесть новых локусов риска болезни Паркинсона.Генетика природы. 2014;46(9):989. пмид:25064009
- 7. Satake W, Nakabayashi Y, Mizuta I, Hirota Y, Ito C, Kubo M, et al. Полногеномное ассоциативное исследование выявляет общие варианты в четырех локусах как генетические факторы риска болезни Паркинсона. Генетика природы. 2009;41(12):1303. пмид:196
- 8. Браак Х., Дель Тредичи К., Руб У., де Вос Р.А., Янсен Стер Э.Н., Браак Э. Стадирование патологии головного мозга, связанной со спорадической болезнью Паркинсона. Нейробиология старения.2003;24(2):197–211. Эпублик 25 декабря 2002 г. пмид:12498954.
- 9. Браак Х., Дель Тредичи К. Приглашенная статья: Патология нервной системы при спорадической болезни Паркинсона. Неврология. 2008;70(20):1916–25. Эпб 2008/05/14. пмид: 18474848.
- 10.
Volpicelli-Daley L, Brundin P. Прионоподобное распространение патологии при болезни Паркинсона. Handb Clin Neurol. 2018; 153:321–35. Эпб 2018/06/12. пмид: 29887143; Центральный PMCID в PubMed: PMC6625652.
- 11.
Варгас Ю.Ю., Грудина С., Зурзоло С.Прионоподобное распространение альфа-синуклеина: от in vitro к in vivo моделям болезни Паркинсона. Aging Res Rev. 2019; 50: 89–101. Эпб 2019/01/29. пмид:306
. - 12. Тофарис Г.К., Гедерт М., Спиллантини М.Г. Трансцеллюлярное размножение и внутриклеточная торговля альфа-синуклеином. Колд Спринг Харб Перспект Мед. 2017;7(9). Эпублик 2016/12/07. пмид: 27920026; Центральный PMCID в PubMed: PMC5580513.
- 13. Jang H, Boltz DA, Webster RG, Smeyne RJ. Вирусный паркинсонизм.Биохимика и биофизика акта. 2009;1792(7):714–21. пмид: 18760350; Центральный PMCID в PubMed: PMC4642437.
- 14. Tulisiak CT, Mercado G, Peelaerts W, Brundin L, Brundin P. Могут ли инфекции вызывать альфа-синуклеинопатии? Prog Mol Biol Transl Sci.
2019; 168: 299–322. Эпублик 2019/11/09. пмид:31699323; Центральный PMCID в PubMed: PMC6857718.- 15. Гармаруди Ф.С., Маршан Д., Хендри Р., Луо Х., Ян Д., Йе С. и др. Репликация вируса Коксаки В3 и патогенез. Будущая микробиология.2015;10(4):629–53. пмид:25865198
- 16. Там ЧП. Вирусный миокардит Коксаки: взаимодействие между вирусом и хозяином в патогенезе сердечных заболеваний. Вирусная иммунология. 2006;19(2):133–46. пмид:16817756
- 17. Мьюир П., Ван Лун А. Энтеровирусные инфекции центральной нервной системы. Интервирусология. 1997;40(2–3):153–66. пмид:9450232
- 18. Чепмен Н.М., Ким К.С. Персистирующая коксакивирусная инфекция: персистенция энтеровируса при хроническом миокардите и дилатационной кардиомиопатии.Курр Топ Микробиол Иммунол. 2008; 323: 275–92. Эпб 2008/03/25. пмид: 18357775.
- 19. Алиджину Э.К., Сане Ф., Энгельманн И., Гинен В., Хобер Д. Устойчивость энтеровирусов как механизм патогенеза диабета 1 типа.
Дисков Мед. 2014;18(100):273–82. Эпублик 27.11.2014. пмид: 25425468.- 20. Доурмашкин Р.Р., МакКолл С.А., Доурмашкин Н., Ханна М.Дж. Вирусоподобные частицы и антиген энтеровируса обнаружены в нейронах ствола мозга при болезни Паркинсона. F1000Исследование.2018;7. пмид:29899977
- 21. Битман Э.Л., Мэсси А., Шивз К.Д., Баррак К.С., Чаманян М., Моррисон Т.Э. и др. Экспрессия альфа-синуклеина ограничивает РНК-вирусные инфекции в головном мозге. Журнал вирусологии. 2016;90(6):2767–82.
- 22. Гейл М. мл., Тан С. Л., Катце М. Г. Трансляционный контроль экспрессии вирусных генов у эукариот. Microbiol Mol Biol Rev. 2000;64(2):239–80. Эпубликовано 06.06.2000. пмид:10839817; Центральный PMCID в PubMed: PMC98994.
- 23. Дунгель П., Канту Ф.М., Молина Дж.А., Ян З.Вирус коровьей оспы как мастер индукции отключения хозяина: нацеливание на процессы центральной догмы и за ее пределами. Возбудители. 2020;9(5):400. пмид:32455727
- 24.
Инглис С. Ингибирование синтеза белка-хозяина и деградация клеточных мРНК при заражении вирусом гриппа и простого герпеса. Молекулярная и клеточная биология. 1982; 2(12):1644. пмид:14582206- 25. Шек Н., Бахенхаймер С.Л. Деградация клеточных мРНК, индуцированная фактором, ассоциированным с вирионами, при заражении вирусом простого герпеса клеток Vero.Журнал вирусологии. 1985; 55(3):601. пмид:4020960
- 26. Яламанчили П., Харрис К., Виммер Э., Дасгупта А. Ингибирование базовой транскрипции полиовирусом: кодируемая вирусом протеаза (3Cpro) ингибирует образование комплекса TBP-TATA box in vitro. Журнал вирусологии. 1996;70(5):2922. пмид:8627767
- 27. Алексопулу Л., Холт А.С., Меджитов Р., Флавелл Р.А. Распознавание двухцепочечной РНК и активация NF-κB Toll-подобным рецептором 3. Природа. 2001;413(6857):732.пмид:11607032
- 28. Kemball CC, Alirezaei M, Flynn CT, Wood MR, Harkins S, Kiosses WB, et al.
Коксакивирусная инфекция индуцирует аутофагоподобные везикулы и мегафагосомы в ацинарных клетках поджелудочной железы in vivo. Журнал вирусологии. 2010;84(23):12110–24. пмид:20861268- 29. Шибутани С.Т., Сайтох Т., Новаг Х., Мюнц С., Йошимори Т. Аутофагия и связанные с аутофагией белки в иммунной системе. Природная иммунология. 2015;16(10):1014–24. пмид:26382870
- 30.Вакабаяси К., Танджи К., Одагири С., Мики Ю., Мори Ф., Такахаши Х. Тельце Леви при болезни Паркинсона и связанных с ней нейродегенеративных расстройствах. Молекулярная нейробиология. 2013;47(2):495–508. пмид:22622968
- 31. Shahmoradian SH, Lewis AJ, Genoud C, Hench J, Moors TE, Navarro PP, et al. Патология Леви при болезни Паркинсона состоит из переполненных органелл и липидных мембран. Неврология природы. 2019;22(7):1099. пмид:31235907
- 32. Ан Дж., Чхве Дж., Джу Ч., Со И, Ким Д., Юн С.И. и др.Восприимчивость клеток первичных корковых нейронов мыши к вирусу Коксаки B.
J Gen Virol. 2004; 85 (часть 6): 1555–64. Эпб 2004/05/29. пмид: 15166439.- 33. Шульц Н.В., Кановка С.С., Тен Эйк Ю.Е., Рыбка В., Судзуки Ю.Ю. Ультраструктурные изменения миоцитов правого желудочка при легочной артериальной гипертензии. Журнал Американской кардиологической ассоциации. 2019;8(5):e011227. пмид:30807241
- 34. Вонг Дж., Чжан Дж., Си С., Гао Г., Мао И., Макманус Б.М. и др. Аутофагосома поддерживает репликацию вируса Коксаки B3 в клетках-хозяевах.Журнал вирусологии. 2008;82(18):9143–53. пмид:18596087
- 35. Клионский Д.Дж., Абдельмохсен К., Абэ А., Абедин М.Дж., Абелиович Х., Асеведо Арозена А. и соавт. Руководство по использованию и интерпретации анализов для мониторинга аутофагии (3-е издание). Аутофагия. 2016;12(1):1–222. Эпб 2016/01/23. пмид: 26799652; Центральный PMCID в PubMed: PMC4835977.
- 36. Йоши С.Р., Мидзусима Н. Мониторинг и измерение аутофагии. Int J Mol Sci. 2017;18(9):1865. Эпб 2017/08/29.
пмид: 28846632; Центральный PMCID в PubMed: PMC5618514.- 37. Xilouri M, Vogiatzi T, Vekrellis K, Park D, Stefanis L. Абберантный α-синуклеин придает токсичность нейронам частично за счет ингибирования аутофагии, опосредованной шаперонами. ПлоС один. 2009;4(5):e5515. пмид:19436756
- 38. Vuorinen T, Kallajoki M, Hyypiä T, Vainionpää R. Острый панкреатит, вызванный вирусом Коксаки B3: анализ гистопатологических и вирусных параметров на мышиной модели. Br J Exp патол. 1989;70(4):395–403. Эпб 1989/08/01. пмид: 2548562; Центральный PMCID в PubMed: PMC2040566.
- 39. Холл Дж. Х., Клеменс Д. Л. Использование вируса Коксаки B3 в качестве модели острого панкреатита. Pancreapedia: База знаний экзокринной поджелудочной железы. 2016.
- 40. Кляйн С, Шлосмахер М.Г. Болезнь Паркинсона, спустя 10 лет после ее генетической революции: несколько ключей к разгадке сложного расстройства. Неврология. 2007;69(22):2093–104. пмид:17761553
- 41.
Джонсон М.Е., Стечер Б., Лабри В., Брундин Л., Брундин П. Триггеры, фасилитаторы и усугубляющие факторы: новое определение патогенеза болезни Паркинсона.Тренды Нейроси. 2019;42(1):4–13. Эпб 2018/10/22. пмид:30342839; Центральный PMCID в PubMed: PMC6623978.- 42. Махул-Мелье А.Л., Бертшер Дж., Махарьян Н., Веренс Л., Круазье М., Каттлер Ф. и др. Процесс образования телец Леви, а не просто фибрилляция альфа-синуклеина, является одним из основных факторов нейродегенерации. Труды Национальной академии наук Соединенных Штатов Америки. 2020;117(9):4971–82. Эпаб 2020/02/23. пмид:32075919.
- 43.Лашуэль ХА. Содержат ли тельца Леви фибриллы альфа-синуклеина? и имеет ли это значение? Краткая история и критический анализ последних сообщений. Нейробиология болезни. 2020:104876. Эпаб 2020/04/28. пмид:32339655.
- 44. Вакабаяши К., Хаяши С., Какита А., Ямада М., Тойосима Ю., Йошимото М. и др. Накопление альфа-синуклеина/NACP является цитопатологическим признаком, общим для болезни телец Леви и множественной системной атрофии.
Акта Нейропатол. 1998;96(5):445–52. Эпублик 26 ноября 1998 г.пмид:9829807.- 45. Katsuse O, Iseki E, Marui W, Kosaka K. Стадии развития корковых телец Леви и их связь с блокировкой аксонального транспорта в головном мозге пациентов с деменцией с тельцами Леви. J Neurol Sci. 2003;211(1–2):29–35. Эпб 2003/05/28. пмид: 12767494.
- 46. Деви Л., Рагхавендран В., Прабху Б.М., Авадхани Н.Г., Анандатиртаварада Х.К. Митохондриальный импорт и накопление альфа-синуклеина нарушают комплекс I в культурах дофаминергических нейронов человека и мозге при болезни Паркинсона.Дж. Биол. Хим. 2008;283(14):9089–100. Эпб 2008/02/05. пмид: 18245082; Центральный PMCID в PubMed: PMC2431021.
- 47. Накамура К., Немани В.М., Азарбал Ф., Скибински Г., Леви Дж.М., Эгами К. и др. Прямая ассоциация с мембраной управляет делением митохондрий белком альфа-синуклеином, ассоциированным с болезнью Паркинсона. Дж. Биол. Хим. 2011;286(23):20710–26. Эпб 2011/04/15.
пмид: 21489994; Центральный PMCID в PubMed: PMC3121472.- 48. Park JH, Burgess JD, Faroqi AH, DeMeo NN, Fiesel FC, Springer W, et al.Альфа-синуклеин-индуцированная митохондриальная дисфункция опосредуется сиртуин-3-зависимым путем. Мол Нейродегенер. 2020;15(1):5. Эпублик 2020/01/15. пмид:31931835; Центральный PMCID в PubMed: PMC6956494.
- 49. Paxinou E, Chen Q, Weisse M, Giasson BI, Norris EH, Rueter SM, et al. Индукция агрегации альфа-синуклеина внутриклеточным нитратным инсультом. Журнал неврологии: официальный журнал Общества неврологии. 2001;21(20):8053–61. Эпублик 2001/10/06. пмид:11588178; Центральный PMCID в PubMed: PMC6763872.
- 50. Vogiatzi T, Xilouri M, Vekrellis K, Stefanis L. α-synuclein дикого типа расщепляется шаперон-опосредованной аутофагией и макроаутофагией в нейрональных клетках. Журнал биологической химии. 2008;283(35):23542–56. пмид:18566453
- 51. Альварес-Эрвити Л., Родригес-Орос М.
С., Купер Дж.М., Кабальеро С., Феррер И., Обесо Дж.А. и др. Маркеры шаперон-опосредованной аутофагии в мозге при болезни Паркинсона. Архив неврологии. 2010;67(12):1464–72. пмид:20697033- 52.Стефанис Л., Эммануилиду Э., Пантазопулу М., Кирик Д., Векреллис К., Тофарис Г.К. Как альфа-синуклеин выводится из клетки? Журнал нейрохимии. 2019;150(5):577–90. Эпб 2019/05/10. пмид:31069800.
- 53. Эйлерсков П., Халтберг Дж. Г., Ван Дж., Карлссон Р., Амбьорн М., Кусс М. и др. Недостаток нейронального IFN-бета-IFNAR вызывает деменцию, подобную болезни Паркинсона и тельцам Леви. Клетка. 2015;163(2):324–39. Эпублик 2015/10/10. пмид: 26451483; Центральный PMCID в PubMed: PMC4601085.
- 54. Нейстат М., Линч Т., Пшедборски С., Холодилов Н., Ржецкая М., Берк Р.Е. Экспрессия альфа-синуклеина в черной субстанции и коре головного мозга при болезни Паркинсона. Мов Беспорядок. 1999;14(3):417–22. Эпубликовано 29 мая 1999 г. пмид:10348463.
- 55. Кингсбери А.
Е., Даниэль С.Е., Сангха Х., Эйзен С., Лис А.Дж., Фостер О.Дж. Изменение экспрессии мРНК альфа-синуклеина при болезни Паркинсона. Мов Беспорядок. 2004;19(2):162–70. Эпб 2004/02/24. пмид: 14978671.- 56. Блишук П.Миокардит у людей и в экспериментальных моделях животных. Front Cardiovasc Med. 2019;6:64. Эпб 2019/06/04. пмид:31157241; Центральный PMCID в PubMed: PMC6532015.
- 57. Feuer R, Ruller CM, An N, Tabor-Godwin JM, Rhoades RE, Maciejewski S, et al. Вирусная персистенция и хроническая иммунопатология в центральной нервной системе взрослых после заражения вирусом Коксаки в неонатальном периоде. Дж Вирол. 2009;83(18):9356–69. Эпб 2009/07/03. пмид: 19570873; Центральный PMCID в PubMed: PMC2738251.
- 58. Рантакаллио П., Джонс П., Моринг Дж., Фон Вендт Л. Связь между инфекциями центральной нервной системы в детстве и шизофренией во взрослом возрасте и другими психозами: 28-летнее наблюдение. Международный журнал эпидемиологии. 1997;26(4):837–43.
пмид:9279617- 59. Вудалл С.Дж., Грэм Д.И. Доказательства локализации энтеровирусных последовательностей в нейронах при болезни двигательных нейронов/боковом амиотрофическом склерозе путем гибридизации in situ. Eur J Histochem.2004;48(2):129–34. Эпб 2004/06/23. пмид: 15208080.
- 60. Xue YC, Ruller CM, Fung G, Mohamud Y, Deng H, Liu H и другие. Энтеровирусная инфекция приводит к трансактивному ответу ДНК-связывающего белка 43 патологии in vivo. Ам Джей Патол. 2018;188(12):2853–62. Эпб 2018/10/03. пмид:30273599.
- 61. Костин С., Пул Л., Эльзассер А., Хайн С., Дрекслер Х.К., Арнон Э. и др. Миоциты умирают по множеству механизмов при отказе человеческого сердца. Исследование тиража. 2003;92(7):715–24.пмид:12649263
- 62. Санбе А., Осинска Х., Саффиц Дж. Э., Глабе К. Г., Кайед Р., Малоян А. и др. Связанная с десмином кардиомиопатия у трансгенных мышей: сердечный амилоидоз. Труды Национальной академии наук. 2004;101(27):10132–6.
пмид:15220483- 63. Вестермарк Г.Т., Крогволд Л., Даль-Йоргенсен К., Людвигссон Дж. Островковый амилоид при недавно начавшемся диабете 1 типа — исследование DiViD. Упсала журнал медицинских наук. 2017;122(3):201–3. пмид:28814132
- 64.Shen WC, Chiu HH, Chow KC, Tsai CH. Результаты МРТ энтеровирусного энцефалоемелита: вспышка на Тайване. AJNR Am J Нейрорадиол. 1999; 20 (10): 1889–95. Эпублик 10 декабря 1999 г. пмид: 10588115.
- 65. Джексон В.Т., Гиддингс Т.Х. младший, Тейлор М.П., Мулиньяве С., Рабинович М., Копито Р.Р. и др. Подрыв клеточного аутофагосомного механизма РНК-вирусами. Биология PLoS. 2005;3(5):e156. пмид:15884975
- 66. Киркегор К., Тейлор М.П., Джексон В.Т. Клеточная аутофагия: капитуляция, избегание и подрывная деятельность микроорганизмов.Природа Обзоры микробиологии. 2004;2(4):301. пмид:15031729
- 67. Yoon SY, Ha YE, Choi JE, Ahn J, Lee H, Kweon HS и др. Вирус Коксаки B4 использует аутофагию для репликации после активации кальпаина в первичных нейронах крыс.
Дж Вирол. 2008;82(23):11976–8. Эпб 2008/09/19. пмид: 18799585; Центральный PMCID в PubMed: PMC2583648.- 68. Саркар С., Олсен А.Л., Сыгнецка К., Лор К.М., Фини М.Б. α-синуклеин нарушает созревание аутофагосом за счет аномальной стабилизации актина.Генетика PLoS. 2021;17(2):e1009359. Эпб 2021/02/09. пмид:33556113; Центральный PMCID в PubMed: PMC7895402.
- 69. Кобаяши С., Орба Ю., Ямагути Х., Такахаши К., Сасаки М., Хасебе Р. и др. Аутофагия ингибирует репликацию вирусного генома и этапы экспрессии генов при инфекции, вызванной вирусом Западного Нила. Исследование вирусов. 2014; 191:83–91. пмид:250
- 70. Сёдзи-Кавата С., Самптер Р., Левено М., Кэмпбелл Г.Р., Зоу З., Кинч Л. и др. Идентификация кандидата терапевтического пептида, индуцирующего аутофагию.Природа. 2013;494(7436):201. пмид:23364696
- 71. Jang H, Boltz D, McClaren J, Pani AK, Smeyne M, Korff A, et al. Воспалительные эффекты высокопатогенной инфекции вируса гриппа H5N1 в ЦНС мышей.
Журнал неврологии. 2012;32(5):1545–59. пмид:22302798- 72. Садасиван С., Занин М., О’Брайен К., Шульц-Черри С., Смейн Р.Дж. Индукция активации микроглии после инфицирования ненейротропным вирусом гриппа A/CA/04/2009 h2N1. ПлоС один.2015;10(4). пмид:25861024
- 73. Jang H, Boltz D, Sturm-Ramirez K, Shepherd KR, Jiao Y, Webster R, et al. Высокопатогенный вирус гриппа H5N1 может проникать в центральную нервную систему и вызывать нейровоспаление и нейродегенерацию. Труды Национальной академии наук. 2009;106(33):14063–8. пмид:19667183
- 74. Маррейрос Р., Мюллер-Шиффманн А., Троссбах С.В., Прикулис И., Хэнш С., Вайдткамп-Питерс С. и др. Нарушение клеточного протеостаза вирусом гриппа А h2N1 вызывает агрегацию α-синуклеина.Труды Национальной академии наук. 2020;117(12):6741–51. пмид:32152117
- 75. Choi YR, Cha SH, Kang SJ, Kim JB, Jou I, Park SM. Прионоподобное распространение альфа-синуклеина регулируется сигнальным путем FcgammaRIIB-SHP-1/2 в нейронах. Cell Rep. 2018;22(1):136–48. Эпублик от 04.01.2018. пмид: 29298416.
- 76. Санджана Н.Е., Шалем О., Чжан Ф. Улучшенные векторы и полногеномные библиотеки для скрининга CRISPR. Природные методы. 2014;11(8):783. пмид:25075903
- 77.Лопачински А.Б., Суитт А.Дж., Смолко К.М., Грей-Гайяр Э., Боргман К.А., Шах М. и соавт. Моделирование полной кинетики вируса Коксаки B3 выявляет человеческие детерминанты обратной связи с клеткой-хозяином. Клеточные системы. 2021;12(4):304–23. е13. пмид:33740397
- 78. Юн С, Пак С, Хан Дж, Кан Дж, Ким Дж, Ли Дж и др. Каспазозависимое высвобождение нуклеосом, связанное со смертью клеток, и молекулярные паттерны, связанные с повреждением. Гибель клеток и болезни. 2014;5(10):e1494–e. пмид:25356863
- 79.Робинсон С.М., Цуэн Г., Син Дж., Мангале В., Рахави С., Макинтайр Л.Л. и др. Вирус Коксаки В выходит из клетки-хозяина в виде сбрасываемых микровезикул, демонстрирующих аутофагосомные маркеры. Возбудители PLoS. 2014;10(4):e1004045. пмид: 24722773
Classici Stranieri – бесплатная, юридическая и бесплатная мультимедийная библиотека. Всеобщее солнце! Мы не Либер, мы свободны.
573 всего просмотров, 4 просмотра сегодня
573 всего просмотров, 4 просмотров сегодня Статическая Википедия: Итальяно – Английский – Французский – Испанский – Тедеско – Португальский – Оландский – Полакко – Руссо – Турко – Шведский – […]
Continua a leggere580 всего просмотров, 2 просмотра сегодня
580 всего просмотров, 2 просмотра сегодня Sono in linea le edizioni 2008 della Static Wikipedia (только текст, niente immagini) на африкаанс и суахили.Potete trovarle ai seguenti […]
Continua a leggere1 421 всего просмотров
1 421 общее количество просмотров Esprimiamo la nostra più Complete e Sentita Solidarietà Agli Amici del Project Gutenberg per l’oscuramento subito sul territorio italiano, su solutione del Tribunale […]
Continua a leggere429 всего просмотров, 2 просмотра сегодня
429 всего просмотров, 2 просмотра сегодня от: BBC News Фотография Стива Лиддьярда маяка Уайтфорд-Пойнт на полуострове Гауэр была названа абсолютным победителем.
Continua a leggere430 всего просмотров, 6 просмотров сегодня
430 всего просмотров, 6 просмотров сегодня от: BBC News Осман Кавала не был осужден, но его задержание поставило турецкого лидера на путь столкновения.
Continua a leggere404 всего просмотра, 2 просмотра сегодня
404 всего просмотров, 2 просмотра сегодня. Создавая свой сериал о Битлз Get Back, режиссер узнал о разочаровании ливерпульской четверки.
Continua a leggere358 всего просмотров, 2 просмотра сегодня
358 всего просмотров, 2 просмотра сегодня. Газеты, опубликованные в четверг, сообщают о гибели 27 человек, направлявшихся в Великобританию через Ла-Манш.
Continua a leggere370 всего просмотров
370 всего просмотров Футболист из Англии станет одним из семи традиционных приглашенных редакторов в праздничный период.
Continua a leggere401 всего просмотров, 2 просмотра сегодня
401 всего просмотров, 2 просмотров сегодня Группа коллег призывает правительство отменить Закон о бродяжничестве, согласно которому сон считается уголовным преступлением […]
Continua a leggere325 всего просмотров, 2 просмотра сегодня
325 всего просмотров, 2 просмотра сегодня Женщины на занятиях крав-мага рассказывают, почему в этом году они ходили на занятия по самообороне.
Continua a leggere335 всего просмотров, 2 просмотра сегодня
335 всего просмотров, 2 просмотра сегодня. Корреспондент Би-би-си Абдужалил Абдурасулов посещает восточную Украину, пока солдаты созерцают военное присутствие России на границе поблизости.
Continua a leggere395 всего просмотров, 2 просмотра сегодня
395 всего просмотров, 2 просмотра сегодня «Манчестер Сити» обыграл «Пари Сен-Жермен», и, поскольку Маурисио Почеттино изо всех сил пытается навязать свой стиль, разговоры о «Манчестер Юнайтед» не пойдут […]
Continua a leggere375 всего просмотров, 2 просмотра сегодня
375 всего просмотров, 2 просмотра сегодня от: BBC News Португальские велосипедные заводы были очень загружены с апреля 2020 года, поскольку популярность велоспорта резко возросла.
Continua a leggere335 всего просмотров, 2 просмотра сегодня
335 всего просмотров, 2 просмотра сегодня Финал Кубка Дэвиса готов перенести в Абу-Даби в рамках пятилетнего контракта, который должен быть утвержден […]
Continua a leggere339 всего просмотров, 2 просмотра сегодня
339 всего просмотров, 2 просмотра сегодня Десятилетия обедов Дэвида Р. Чана в 8 000 китайских закусочных научили его об Америке и о себе.из: BBC News
Continua a leggere353 всего просмотров, 2 просмотра сегодня
353 всего просмотров, 2 просмотра сегодня. Матч разминки внутри сборной Англии «Эшес» в Брисбене прерван без подачи мяча в последний день из-за дождя. […]
Continua a leggere437 всего просмотров, 6 просмотров сегодня
437 всего просмотров, 6 просмотров сегодня Ахмад Арбери погиб в результате «современного линчевания» — местные жители говорят, что призрак расизма присутствует всегда.из: BBC News
Continua a leggere322 всего просмотров
322 всего просмотров «Реал Мадрид» выходит в плей-офф Лиги чемпионов 25-й год подряд, одержав комфортную победу над тираспольским «Шерифом». из: BBC News
Continua a leggere372 всего просмотра, 2 просмотра сегодня
372 всего просмотров, 2 просмотра сегодня Энергетическая компания с 1,7 млн клиентов переведена на особую форму управления.
Continua a leggere380 всего просмотров, 6 просмотров сегодня
380 всего просмотров, 6 просмотров сегодня. Английскому футболу нужен независимый регулятор, чтобы остановить его «шатание от кризиса к кризису», — говорит председатель обзора, проводимого фанатами […]
Continua a leggere126 089 всего просмотров, 161 просмотр сегодня
126 089 всего просмотров, 161 просмотр сегодня от: BBC News «Манчестер Сити» выходит в 1/8 финала Лиги чемпионов как победитель группы, выходя сзади […]
Continua a leggereВирус Коксаки – Википедия – Enzyklopädie
Virus, das Verdauungsstörungen und manchmal Herzschäden verursacht
CoxSackievireen Sind Einige Verwandte Enterovireen, Die Zu Den Gehören Picornaviridae Familie Nicht Umhüllter, Linearer, Einzelsträngiger РНК-Вирен Mit Positivem Sinn Sowie Deren Gattung Enterovirus , Zu denen Auch Poliovirus und Echovirus Gehören.Enteroviren gehören zu den häufigsten und wichtigsten humanpathogenen Erregern, und normalerweise werden ihre Mitglieder auf fäkal-oralem Weg übertragen. Coxsackieviren haben viele Eigenschaften mit Poliovirus gemeinsam. Bei der Bekämpfung von Poliovirus-Infektionen in weiten Teilen der Welt wurde mehr Aufmerksamkeit auf das Verständnis der Nicht-Polio-Enteroviren wie Coxsackievirus gerichtet.
Coxsackieviren gehören zu den Hauptursachen für aseptische Meningitis (die anderen üblichen Verdächtigen sind Echovirus und Pumsvirus).
Eintritt des Coxsackievirus in Zellen, insbesondere in Endothelzellen, wird durch Coxsackievirus und Adenovirusrezeptor vermittelt.
Coxsackieviren werden werden basierend auf frühen Beobachtungen ihrer Pathogenität bei neugeborenen Mäusen in Viren der Gruppen A und B unterteilt. [1] ES Wurde Fest gruppe, DASS COXSACKEVIREN DER GRUPPE EINE SCHLAFFE Lähmung Verurursachen (Die Durch Generalisierte MyoStisite Veroursacht Wurde), Während Coxsackeviren der Gruppe B Eine Spastische Lähmung Verurursachen (Aufgrund Einer Fokalen Muskelverletzung und Eill Degeneration des Neurollen Gewebes).Eswerden mindestens 23 Serotypen (1–22, 24) der Gruppe A und sechs Serotypen (1–6) der Gruppe Berkannt.
Группа А[править]
Im Allgemeinen neigen Coxsackieviren der Gruppe A dazu, die Haut und die Schleimhäute zu infizieren und Herpangina zu verursachen. острый геморрагический конъюнктивит; und Hand-, Fuß- und Mundkrankheit (HFM). [2]
Sowohl Coxsackieviren der Gruppe A als auch der Gruppe B können unspezifische fieberhafte Erkrankungen, Hautausschläge, Erkrankungen der oberen Atemwege und aseptische Meningitis verursachen.
Die Grundreproductionszahl (R0) for Coxsackievirus A16 (Cox A16) wurde auf einen Median von 2,50 mit einem Interquartilbereich 1,96 bis 3,67 geschätzt. [3]
Группа B.[править]
Coxsackieviren der Gruppe B neigen dazu, Herz, Pleura, Bauchspeichelmrüse und Leber zu infizieren und Pleurodynie, Myokarditis, Perikarditis und Hepatitis (Entzündung der Leber, die nicht mit den hepatotropen Viren zusammenhängt) zu verursachen. Eine Coxsackie B-Infektion des Herzens kann zu einem Perikarderguss führen.
Die Entwicklung von инсулиноподобный диабет (IDDM) wurde kürzlich mit einer kürzlich aufgetretenen enteroviralen Infektion in Verbindung gebracht, insbesondere mit einer Coxsackievirus B-панкреатит. Diese Beziehung wird derzeit weiter untersucht.
Das Sjögren-Syndrom wird ab Januar 2010 auch im Zusammenhang mit dem Coxsackievirus untersucht. [4]
Таксономия[править]
Es gab 29 Arten von Coxsackieviren bis 1999, als zwei von ihnen abgeschafft wurden und der Rest mit anderen Arten verschmolz. [5]
Geschichte[править]
Die Coxsackieviren wurden 1948–49 entdeckt von Gilbert Dalldorf, einem Wissenschaftler, der am New Yorker Gesundheitsministerium в Олбани, Нью-Йорк, arbeitet.
Dalldorf in Zusammenarbeit mit Grace Sickles, [11] [12] hatte nach einem Heilmittel für Poliomyelitis gesucht. Frühere Arbeiten, die Dalldorf an Affen durchgeführt hatte, deuteten darauf hin, dass aus einem Nicht-Polio-Virus-Präparat gewonnene Flüssigkeit vor den lähmenden Auswirkungen von Polio schützen könnte.Mit neugeborenen Mäusen als Vehikel versuchte Dalldorf, solche Schutzviren aus dem Kot von Polio-Patienten zu isolieren. Bei der Durchführung dieser Experimente entdeckte er Viren, die häufigmilde oder nichtparalytische Kinderlähmung imitierten. Die Virusfamilie, die er entdeckte, erhielt schließlich den Namen Coxsackie aus Coxsackie, New York, einer kleinen Stadt am Hudson River, in der Dalldorf die ersten Stuhlproben erhalten hatte. [13]
Dalldorf arbeitete auch mit Gifford an vielen frühen Veröffentlichungen zusammen. [14] [15] [16] [17]
Anschließend wurde festgestellt, dass die Coxsackieviren eine Vielzahl von Infektionen verursachen, einschließlich epidemischer Pleurodynie (Bornholm-Krankheit), und sie wurden aufgrund ihrer Pathologie bei neugeborenen Mäusen in die Gruppen A und B unterteilt. (Coxsackie A-Virus verursacht Lähmungen und Tod der Mäuse mit ausgedehnter Skelettmuskelnekrose; Coxsackie B verursacht weniger schwere Infektionen bei den Mäusen, jedoch mit Schäden an mehr Organsystemen wie Herz, Gehirn, Leber, Bauchspeicheldrüse und Skelettmuskeln.)
Die Verwendung von Säuglingsmäusen war nicht Dalldorfs Idee, sondern wurde ihm in einem Artikel der dänischen Wissenschaftler Orskov und Andersen aus dem Jahr 1947 zur Kenntnis gebracht, die solche Mäuse zur Untersuchung eines Mausvirus verwendeten. Die Entdeckung der Coxsackieviren regte viele Virologen zur Verwendung dieses Systems and führte letztendlich zur Isolierung einer großen Anzahl sogenannter «enterischer» Viren aus dem Magen-Darm-Trakt, die nicht mit Poliovirus in Zusammenhang standen und von denen einige onkogen (Krebs-waren) верурсачен).
Die Entdeckung der Coxsackieviren ergab weitere Hinweise darauf, dass Viren manchmal das Wachstum und die Replikation eines anderen Wirtstiers gegenseitig stören können. Andere Forscher fanden heraus, dass diese Störung durch eine vom Wirtstier produzierte Substanz vermittelt werden kann, ein Protein, das jetzt als Interferon bekannt ist. Интерферон шляпа seitdem eine herausragende Rolle bei der Behandlung einer Vielzahl von Krebsarten und Infektionskrankheiten gespielt. EIN C F G H J J K «Geschichte der ictv-taxonomie: Enterovirus C. EIN C G h ich «Geschichte der ICTV-Taxonomie: Enterovirus A. «». Международный комитет по таксономии фон Вирен (ICTV) . EIN C G «Geschichte der ICTV-Taxonomie: Enterovirus B. Далльдорф Г., Сиклз Г.М. (июль 1948 г.). «China sagt, es kontrolliert den Virusausbruch bei Kindern». Рейтер . 19 мая 2007 г. .
Внешние ссылки[править]
Erkrankungen der Haut und der Gliedmaßen nach Morphologie
Вахстум Hautausschläge Mit
эпидермальный
BeteiligungOhne
эпидермальный
BeteiligungВершиденес
ШтёрунгенИнфекции полости рта и другие распространенные поражения | Секретарь педиатрии
Язвы во рту у детей
Язва во рту (в данном случае связанная с афтозным стоматитом) на слизистой оболочке губ (слизистая оболочка нижней губы)
https://en.wikipedia.org/wiki/Mouth_ulcer
Герпетический гингивостоматит ВПГ 1 и ВПГ 2Передача происходит при прямом контакте с поражениями или оральными выделениями. Чаще встречается у детей 1-3 лет. Проявления в полости рта зависят от того, является ли инфекция первичной или рецидивирующей.
Первичное заражение происходит примерно через 1 неделю после контакта с инфицированным человеком.
- может быть субклиническим
- может быть связан с продромальным периодом лихорадки, недомогания и раздражительности
- оральная инфекция, проявляющаяся красными отечными деснами и скоплениями мелких пузырьков.Появление может быть внезапным с началом снижения потребления перорального приема, боли, лихорадки, слюнотечения и неприятного запаха изо рта.
- Могут быть вовлечены все поверхности полости рта, везикулы появляются в течение 3-5 дней
- везикулы могут сливаться и разрываться, образуя большие болезненные язвы в тканях полости рта и периоральной области. Сероватая пленка на желтом основании.
- Может сопровождаться лихорадкой, артралгией, головной болью и шейной лимфаденопатией
- периоральные язвы могут быть обнаружены у большинства детей через несколько дней после появления поражений в полости рта.Эти периоральные поражения помогают дифференцировать ВПГ-инфекцию от афтозных язв, герпетической ангины и болезни рук, ящура .
Рецидивирующее заболевание : После первичной инфекции ВПГ мигрирует в узел тройничного нерва, где становится латентным.
- реактивации может предшествовать воздействие солнечного света, холода, травмы, стресса или иммуносупрессии
- может иметь начало в течение нескольких дней с продромальным зудом, жжением или легким дискомфортом.Язвы гораздо менее болезненны, чем первичное заболевание, и чаще всего обнаруживаются на губах.
Диагностика
- Мазок по Цанку – мазок из везикул на предметное стекло и окрашивание по Гейсме или Райту. Увидим многоядерные гигантские клетки
- Культура клеток — результаты через 1-3 дня
- Прямой флуоресцентный тест на антитела (DFA)
- Серологическое исследование
- ПЦР
Лечение
- Поддерживающее лечение с использованием анальгетиков, фруктового мороженого, отказом от цитрусовых или острой пищи и полосканием глиоксидом для гигиены полости рта.
- Вязкий лидокаин (ксилокаин) можно наносить перед едой, чтобы обеспечить пероральный прием. Будьте осторожны с маленькими детьми, его следует наносить с помощью ватного аппликатора только 3-4 раза в день. Дайте указания родителям не давать большие суммы.
- Половина смеси Маалокса и дифенилгидрамина, нанесенная на язвы, может помочь облегчить симптомы.
- Местные противовирусные препараты не работают.
- Использование перорального ацикловира — если его использовать в течение 48-72 часов, может сократить продолжительность симптомов и выделения.
Осложнения
- хозяевам с ослабленным иммунитетом может потребоваться ацикловир.
- аутоинокуляция может привести к образованию везикул на других поверхностях слизистых оболочек, в глазах и образованию герпетического панариция на пальцах.
Губной герпес нижней губы. Обратите внимание на волдыри в группе, отмеченной стрелкой.
https://en.wikipedia.org/wiki/Herpes_simplex
Заболевания рук, ящура и рта
Герпетический панариций на пальце
https://en.wikipedia.org/wiki/Herpes_simplex- Обычно возникают летом и осенью и вызываются инфекциями энтеровирусами Коксаки А16, наиболее часто.
- Передача фекально-оральная, реже орально-оральная. Вирус может распространяться в течение нескольких месяцев после заражения.
- Часто встречается у малышей и детей школьного возраста.
- 1-2 дня лихорадки и боли в горле с последующим появлением везикул на слизистой оболочке щек и языке.Высыпания слегка болезненны. Везикулы также обнаруживаются на ладонях, подошвах, конечностях и в области ягодиц. Генитальные поражения встречаются редко.
- Дети, как правило, менее болезненны, чем дети с герпетическим гингивостоматитом.
- Рассасывается в течение 2-3 дней без осложнений, лечение поддерживающее.
Герпангина
- В основном вызывается вирусами Коксаки группы А, а также вирусами Коксаки группы В и эховирусами.
- Наибольшая заболеваемость летом и ранней осенью в возрастной группе 3-10 лет
- Продромальная лихорадка, боль в горле и дисфагия могут предшествовать на 1-2 дня развитию экзантемы.
- Серые везикулы размером с булавочную головку вскрываются, образуя большие язвы, покрытые фибрином. Обычно на передних небных дужках, задней части глотки и мягком небе. В среднем 5 поражений, хотя в некоторых случаях их может быть до 16.
- в дополнение к лихорадке и поражениям полости рта могут возникать головная боль, рвота и боль в животе.
- Инфекция самокупируется, симптомы обычно легкие, а поражения исчезают в течение 3-5 дней. Снижение потребления PO может привести к обезвоживанию.
- Лечение поддерживающее и направлено на уменьшение боли.
- Ветряная оспа может иметь везикулы во рту, но диффузные кожные проявления и образование корок на везикулах помогают отличить ее от других инфекций.
- Острый псевдомембранозный кандидоз, или молочница, является распространенной местной инфекцией, вторичной по отношению к чрезмерному росту грибка Candida Albicans.
- Возникает у детей после попадания во влагалище, у детей, получающих антибиотики, химиотерапию, облучение, ингаляционные глюкокортикоиды, а также у детей с ослабленным иммунитетом.
- Псевдомембранозная форма наиболее распространена и характеризуется белыми бляшками на слизистой оболочке щек, языке, небе и ротоглотке.
- при протирании может оставлять красную, шероховатую и болезненную поверхность
- Также может проявляться ангулярным хейлитом или перлешем.
- У пациентов с ВИЧ (-) лечение состоит в следующем:
- Нистатин взмахнуть и проглотить
- Если нет ответа, пероральный флуконазол 100-200 мг в день
- Рецидивы распространены у пациентов с немодифицируемыми факторами риска.Людям с персистирующей молочницей можно профилактически принимать флуконазол в дозе 100 мг в день.
- У ВИЧ-инфицированных (+) клинические проявления могут быть более разнообразными и тяжелыми. Молочница является наиболее распространенной оппортунистической инфекцией у лиц с ВИЧ
. Лечение, как правило, более агрессивное, чтобы предотвратить инфекцию пищевода и осложнения.
Аптозные язвы
Кандидоз полости рта у младенцев. В очень молодом возрасте иммунная система еще не полностью сформировалась.
https://en.wikipedia.org/wiki/Oral_candidiasis- Этиология неясна, хотя связана со многими основными состояниями. Не вторично по отношению к ВПГ-инфекции.
- Стресс, гормональные факторы, инфекции, пищевая гиперчувствительность, иммунные нарушения, семейная предрасположенность — все это связано с возникновением
- Лаурилсульфат натрия, обнаруженный во многих зубных пастах, был связан со вспышками.
- Дефицит витаминов и минералов может способствовать
- Чаще встречается в детском и подростковом возрасте
- Большие изъязвления с серым фибринозным экссудатом на слизистой оболочке губ, щек или языка.Окруженный красным ореолом. Вылечить за 10-14 дней
- Очень больно
- Лечение направлено на облегчение симптомов
- Нанесение пасты/мази на язвы с триамцинолона ацетонидом, флуоцинонидом и амлексаноксом. Orabase — один из вариантов. Раннее применение может привести к ускоренному заживлению.
- Коррекция дефицита витаминов и минералов
- Доступны специальные зубные пасты без лаурилсульфата натрия.
- Травматические язвы являются наиболее распространенными язвами полости рта у детей.
- после механической, химической или термической травмы.
- Бег и падение с инородным телом во рту может привести к «язвенному» поражению
- Поражения обычно заживают в течение 2 недель.
- Хронически кусающие щеки и другие лица с «самооскорбительным» поведением могут потребовать более комплексного лечения, включая защиту от укусов, накладки на губы, извлечение и седацию.
- Рецидивирующие язвы во рту и в области гениталий, лихорадка и воспаление глаз.
- Язвы во рту болезненны.
- Ассоциируется с артритом, воспалительным заболеванием кишечника, аномалиями ЦНС и кожными проявлениями.
- Андреэ М. Как распознать инфекцию вируса простого герпеса 1 типа и справиться с ней. Современная педиатрия Февраль 2004 г.
- Хадсон Б., Пауэлл К. На пути к доказательной медицине для педиатров. Улучшает ли пероральный прием ацикловира клинические исходы у иммунокомпетентных детей с первичным гингивостоматитом, вызванным простым герпесом? Arch Dis Child, февраль 2009 г.
- Скалли К. Аптозное изъязвление. NEJM 13 июля 2006 г.
- Крол Д., Абд Килс М. Заболевания полости рта. Обзор педиатрии, январь 2007 г.
- Делани Дж., Килс М. Педиатрическая оральная патология. Заболевания мягких тканей и пародонта. Педиатрические клиники Северной Америки, октябрь 2000 г. .
Вернуться к оглавлению
Школы столичного региона справляются со вспышкой вируса Коксаки
Пять школ в Большом столичном регионе сообщили о случаях заболевания «руки, ящур и рот», также известного как вирус Коксаки.
Центры по контролю и профилактике заболеваний говорят, что вирусы Коксаки являются частью того же семейства вирусов, которые вызывают полиомиелит и гепатит А и живут в пищеварительном тракте человека.
Д-р Алан Сандерс, главный врач программы St. Peter’s Health Partners: «Таким образом, профилактики вируса Коксаки на самом деле не существует, он появляется каждый год, чаще в конце лета, начале осени. Он легко передается от человека к человеку, так что очевидно если у кого-то это диагностировано, это очевидно, часто это начинается с сыпи на ногах, на руках и небольших поражений во рту.»
Это заболевание обычно поражает детей в возрасте до пяти лет. Но взрослые также могут заразиться, о чем свидетельствует ряд случаев среди игроков Высшей бейсбольной лиги в этом году. Сандерс говорит, что обычно это не представляет серьезной угрозы для здоровья. в редких случаях может привести к более серьезным проблемам с воспалением вокруг сердца. Это могло бы быть немного более обеспокоено даже формой небактериального или того, что мы называем вирусным менингитом, который обычно полностью бессимптомен через пару дней.И что важно, для этого вируса нет противовирусных препаратов, так что он действительно может нанести ущерб людям в течение нескольких дней, но вы ничего не можете сделать с А, предотвратить его или Б вылечить, он должен идет своим естественным путем», — сказал Сандерс.
Представители органов здравоохранения Лонг-Айленда сообщили о множестве случаев в конце июля и начале августа, и, похоже, вирус переместился на север штата. Впервые он появился в округе Монтгомери в Центральной школе Фонда-Фултонвилля. District, затем появился в Центральном школьном округе Бродалбин-Перт и Школьном округе Большого Амстердама.
Официальные лица приняли меры предосторожности, отменив запланированный на пятницу вечерний футбольный матч между средней школой Амстердама и средней школой Бродалбин-Перт на прошлой неделе. Школьный округ Cohoes и расширенный школьный округ Gloversville были последними, кто сообщил о наличии вируса среди учащихся в конце прошлой недели.
Представитель округа Кохуз Аарон Кегвин говорил с WAMC в понедельник. «На прошлой неделе у нас было четыре подтвержденных случая вируса Коксаки в одной из наших начальных школ, это начальная школа Абрахама Лансинга.И я только что разговаривал с нашим директором, и с тех пор у нас был еще один подтвержденный случай, так что до пяти. Мы просто сообщаем всем нашим родителям, каковы симптомы, и что если у их детей возникнут какие-либо из этих симптомов, они должны обратиться к своему врачу.
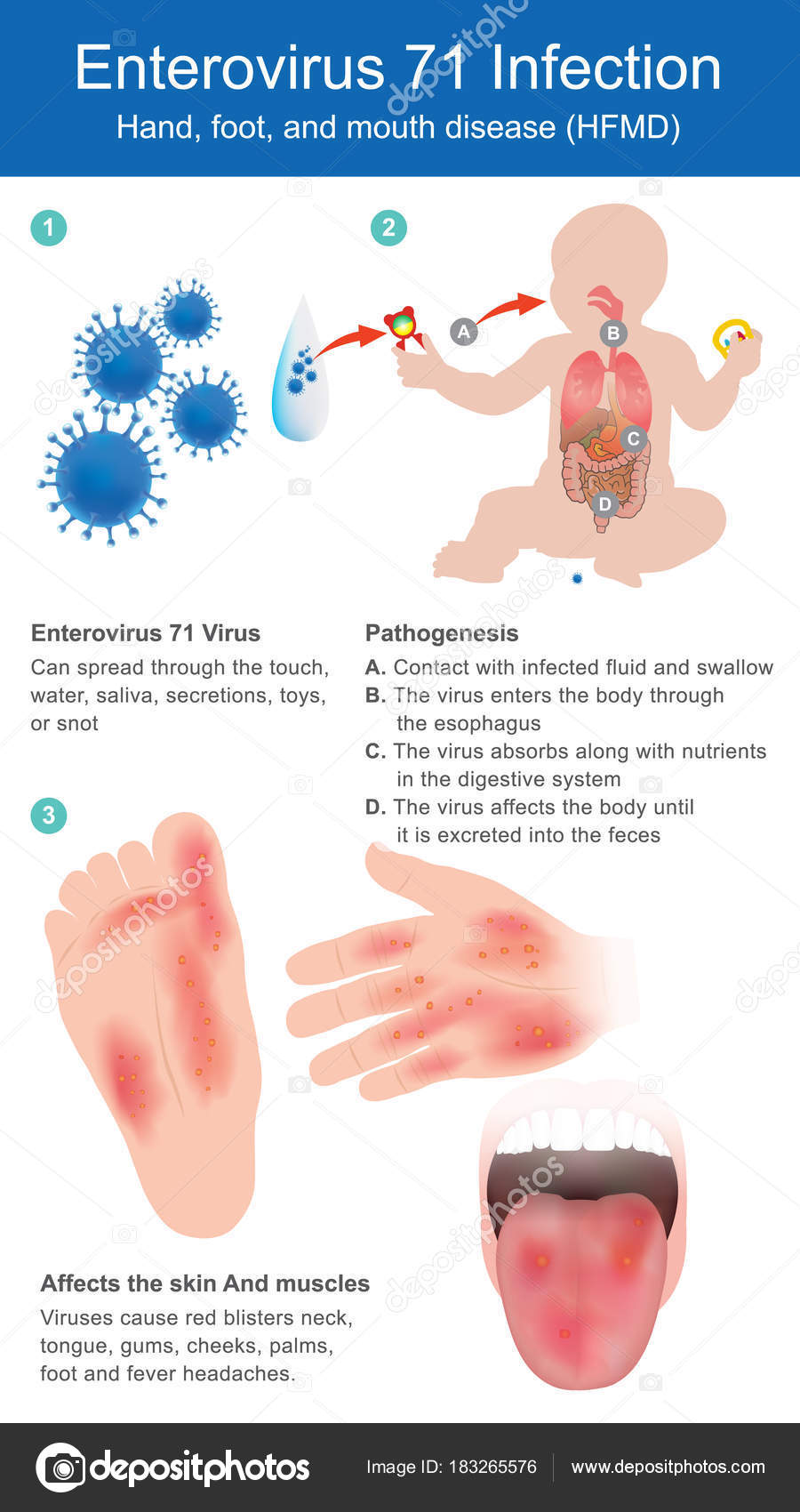 , Лилль К.М., До К.Б., Эрнандес Д.Г., Саад М. и соавт. Крупномасштабный метаанализ данных об ассоциации всего генома выявил шесть новых локусов риска болезни Паркинсона.Генетика природы. 2014;46(9):989. пмид:25064009
, Лилль К.М., До К.Б., Эрнандес Д.Г., Саад М. и соавт. Крупномасштабный метаанализ данных об ассоциации всего генома выявил шесть новых локусов риска болезни Паркинсона.Генетика природы. 2014;46(9):989. пмид:25064009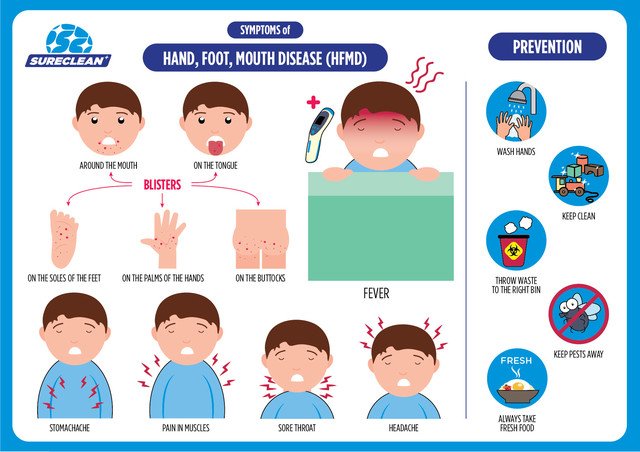 Прионоподобное распространение патологии при болезни Паркинсона. Handb Clin Neurol. 2018; 153:321–35. Эпб 2018/06/12. пмид: 29887143; Центральный PMCID в PubMed: PMC6625652.
Прионоподобное распространение патологии при болезни Паркинсона. Handb Clin Neurol. 2018; 153:321–35. Эпб 2018/06/12. пмид: 29887143; Центральный PMCID в PubMed: PMC6625652. 2019; 168: 299–322. Эпублик 2019/11/09. пмид:31699323; Центральный PMCID в PubMed: PMC6857718.
2019; 168: 299–322. Эпублик 2019/11/09. пмид:31699323; Центральный PMCID в PubMed: PMC6857718. Дисков Мед. 2014;18(100):273–82. Эпублик 27.11.2014. пмид: 25425468.
Дисков Мед. 2014;18(100):273–82. Эпублик 27.11.2014. пмид: 25425468. Инглис С. Ингибирование синтеза белка-хозяина и деградация клеточных мРНК при заражении вирусом гриппа и простого герпеса. Молекулярная и клеточная биология. 1982; 2(12):1644. пмид:14582206
Инглис С. Ингибирование синтеза белка-хозяина и деградация клеточных мРНК при заражении вирусом гриппа и простого герпеса. Молекулярная и клеточная биология. 1982; 2(12):1644. пмид:14582206 Коксакивирусная инфекция индуцирует аутофагоподобные везикулы и мегафагосомы в ацинарных клетках поджелудочной железы in vivo. Журнал вирусологии. 2010;84(23):12110–24. пмид:20861268
Коксакивирусная инфекция индуцирует аутофагоподобные везикулы и мегафагосомы в ацинарных клетках поджелудочной железы in vivo. Журнал вирусологии. 2010;84(23):12110–24. пмид:20861268 J Gen Virol. 2004; 85 (часть 6): 1555–64. Эпб 2004/05/29. пмид: 15166439.
J Gen Virol. 2004; 85 (часть 6): 1555–64. Эпб 2004/05/29. пмид: 15166439. пмид: 28846632; Центральный PMCID в PubMed: PMC5618514.
пмид: 28846632; Центральный PMCID в PubMed: PMC5618514. Джонсон М.Е., Стечер Б., Лабри В., Брундин Л., Брундин П. Триггеры, фасилитаторы и усугубляющие факторы: новое определение патогенеза болезни Паркинсона.Тренды Нейроси. 2019;42(1):4–13. Эпб 2018/10/22. пмид:30342839; Центральный PMCID в PubMed: PMC6623978.
Джонсон М.Е., Стечер Б., Лабри В., Брундин Л., Брундин П. Триггеры, фасилитаторы и усугубляющие факторы: новое определение патогенеза болезни Паркинсона.Тренды Нейроси. 2019;42(1):4–13. Эпб 2018/10/22. пмид:30342839; Центральный PMCID в PubMed: PMC6623978.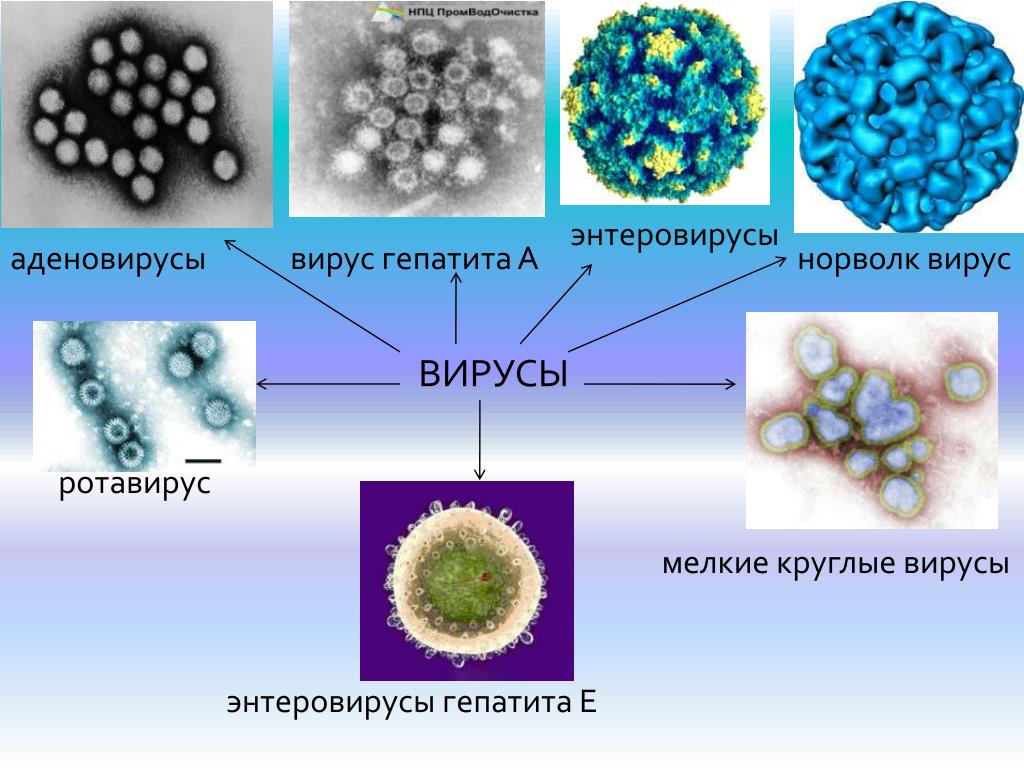 Акта Нейропатол. 1998;96(5):445–52. Эпублик 26 ноября 1998 г.пмид:9829807.
Акта Нейропатол. 1998;96(5):445–52. Эпублик 26 ноября 1998 г.пмид:9829807. пмид: 21489994; Центральный PMCID в PubMed: PMC3121472.
пмид: 21489994; Центральный PMCID в PubMed: PMC3121472. С., Купер Дж.М., Кабальеро С., Феррер И., Обесо Дж.А. и др. Маркеры шаперон-опосредованной аутофагии в мозге при болезни Паркинсона. Архив неврологии. 2010;67(12):1464–72. пмид:20697033
С., Купер Дж.М., Кабальеро С., Феррер И., Обесо Дж.А. и др. Маркеры шаперон-опосредованной аутофагии в мозге при болезни Паркинсона. Архив неврологии. 2010;67(12):1464–72. пмид:20697033 Е., Даниэль С.Е., Сангха Х., Эйзен С., Лис А.Дж., Фостер О.Дж. Изменение экспрессии мРНК альфа-синуклеина при болезни Паркинсона. Мов Беспорядок. 2004;19(2):162–70. Эпб 2004/02/24. пмид: 14978671.
Е., Даниэль С.Е., Сангха Х., Эйзен С., Лис А.Дж., Фостер О.Дж. Изменение экспрессии мРНК альфа-синуклеина при болезни Паркинсона. Мов Беспорядок. 2004;19(2):162–70. Эпб 2004/02/24. пмид: 14978671. пмид:9279617
пмид:9279617 пмид:15220483
пмид:15220483 Дж Вирол. 2008;82(23):11976–8. Эпб 2008/09/19. пмид: 18799585; Центральный PMCID в PubMed: PMC2583648.
Дж Вирол. 2008;82(23):11976–8. Эпб 2008/09/19. пмид: 18799585; Центральный PMCID в PubMed: PMC2583648. Журнал неврологии. 2012;32(5):1545–59. пмид:22302798
Журнал неврологии. 2012;32(5):1545–59. пмид:22302798