Почему женщинам намного тяжелее рожать, чем их далеким предкам?
- Хелен Бриггс
- Би-би-си
Автор фото, Getty Images
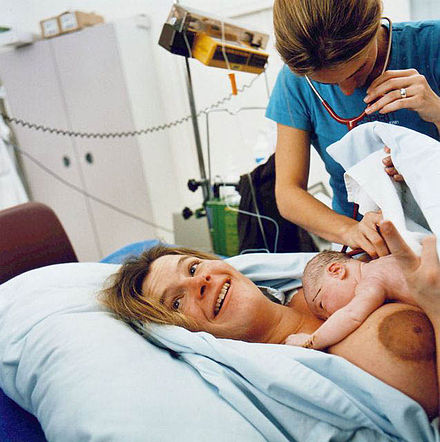
Роды у современных женщин, как правило, проходят мучительно и длятся довольно долго, иногда не один день, а роженице нередко бывает необходима помощь.
Так почему у ближайших биологических родственников человека, например, шимпанзе, роды происходят легче, они длятся лишь несколько часов и роженице не требуется ничья помощь?
Чтобы ответить на этот вопрос, ученые попробовали выяснить, как рожали древнейшие предки человека.
Два миллиона лет назад предкам людей рожать было относительно легко. Это выяснилось в ходе исследования древних окаменелостей.
Для австралопитека седиба, жившего 1,95 млн лет назад на территории нынешней ЮАР, процесс родов был относительно легким, рассказала участник исследования антрополог Натали Лаудицина.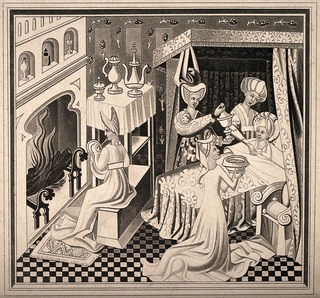
«У головы и плеч плода было достаточно места для прохождения даже самых тесных материнских родовых каналов», — сказала она.
В наши дни ситуация изменилась. Из-за размера и формы современного таза человека (компромисс, необходимый для ходьбы в вертикальном положении) и большого размера головы ребенка плод выходит с трудом.
Во время родов младенцу приходится несколько раз повернуться в родовом канале.
Изучая тазовые кости наших древних человекоподобных родственников, ученые примерно могут понять, какими были роды в основе генеалогического древа человека.
Автор фото, Laudicina et al, 2019
Подпись к фото,Реконструкция родов австралопитека седиба
Но нельзя сказать, что роды неизменно становились труднее по мере эволюции человека.
По словам Лаудицины, при изучении окаменелостей самки австралопитека афарского, названной Люси, выяснилось, что ее роды проходили труднее, чем у австралопитека седиба, потому что в ее случае плоду было более тесно в родовом канале.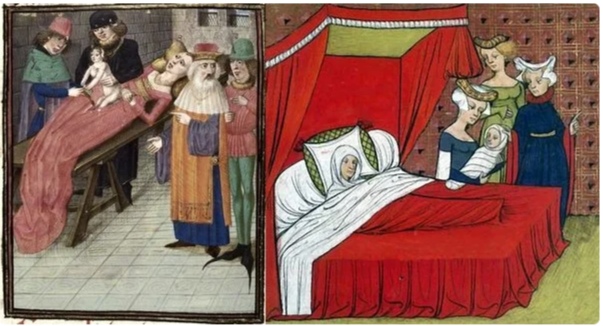
При этом австралопитек афарский жил на миллион лет раньше.
«Многие полагают, что эволюция родов — это переход от легких у обезьян к трудным в наши дни. Но мы видим, что это не так», — рассказала Лаудицина.
По ее словам, сложно определить, когда роды стали трудными и перешли в современную фазу, потому что каждая окаменелость из генеалогического древа человека демонстрирует свои родовые проблемы.
И даже в наши дни роды у женщин бывают разными: у некоторых роды проходят относительно легко и быстро, другие рожают в муках более 20 часов.
Роды- история и старинные картинки | Блогер prodeus на сайте SPLETNIK.RU 18 февраля 2012
Роды, 1800.
Уже в самых древнейших письменных памятниках человеческой истории, в священных книгах индусов, египтян и евреев упоминается об акушерках как особом классе специалистов, а у древних греков и римлян многие богини почитались как покровительницы рожениц
Египетский демон Bes- защитник рожениц и детей. У разных народов древнего мира познания в акушерстве были различные, отсюда различен и характер помощи. Так, у египтян, евреев и китайцев акушерская помощь находилась всецело в руках женщин (повитух). У древних египтян существовало особое сословие женщин, оказывавших пособие роженицам. Каковы были их знания, можно судить по тому, как они диагностировали беременность. Чтобы узнать, беременна ли женщина, ей давали напиток, приготовленный из особой травы (буду-ду-ка), и молока женщины, родившей мальчика. Если этот напиток вызывал рвоту, значит женщина беременна, в противном случае беременность отсутствовала.
Довольно своеобразным методом определялся и пол будущего ребенка. Для этого брали зерна ячменя и пшеницы, смачивали их мочой беременной женщины и следили за прорастанием семян. Если первой прорастала пшеница, то предсказывали, что будет девочка, если ячмень — мальчик. Если родившийся ребенок был очень криклив (причины не доискивались), то для успокоения ему давали смесь из семян мака и мышиного помета.
У разных народов древнего мира познания в акушерстве были различные, отсюда различен и характер помощи. Так, у египтян, евреев и китайцев акушерская помощь находилась всецело в руках женщин (повитух). У древних египтян существовало особое сословие женщин, оказывавших пособие роженицам. Каковы были их знания, можно судить по тому, как они диагностировали беременность. Чтобы узнать, беременна ли женщина, ей давали напиток, приготовленный из особой травы (буду-ду-ка), и молока женщины, родившей мальчика. Если этот напиток вызывал рвоту, значит женщина беременна, в противном случае беременность отсутствовала.
Довольно своеобразным методом определялся и пол будущего ребенка. Для этого брали зерна ячменя и пшеницы, смачивали их мочой беременной женщины и следили за прорастанием семян. Если первой прорастала пшеница, то предсказывали, что будет девочка, если ячмень — мальчик. Если родившийся ребенок был очень криклив (причины не доискивались), то для успокоения ему давали смесь из семян мака и мышиного помета.

 Акушерская помощь ими оказывалась только в случаях тяжелых родов. Им были известны некоторые хирургические методы родоразрешения, в частности, они знали о кесаревом сечении, которое на живых в то время не производилось. Об этой операции на мертвой женщине с целью извлечения живого ребенка повествует и древнегреческий миф о рождении самого бога медицины Асклепия, который был извлечен из трупа матери отцом своим Аполлоном.
Оказанием помощи в родах в Древней Греции занимались исключительно женщины, которых греки называли «перерезывателями пуповины» («omphalotomoi»). Если роды бывали трудные и акушерка (повитуха) видела, что самостоятельно помощь оказать не может, она обращалась как это было и в Индии к врачу-мужчине. Деятельность греческих акушерок была довольно разнообразной: они не только оказывали помощь при родах и в послеродовом периоде, по занимались и прерыванием беременности. В Древней Греции прерывание беременности в ранних сроках не преследовалось. Эту операцию допускал знаменитый древнегреческий философ и естествоиспытатель Аристотель, считая, что в ранний период развития беременности плод не имеет еще сознания.
Акушерская помощь ими оказывалась только в случаях тяжелых родов. Им были известны некоторые хирургические методы родоразрешения, в частности, они знали о кесаревом сечении, которое на живых в то время не производилось. Об этой операции на мертвой женщине с целью извлечения живого ребенка повествует и древнегреческий миф о рождении самого бога медицины Асклепия, который был извлечен из трупа матери отцом своим Аполлоном.
Оказанием помощи в родах в Древней Греции занимались исключительно женщины, которых греки называли «перерезывателями пуповины» («omphalotomoi»). Если роды бывали трудные и акушерка (повитуха) видела, что самостоятельно помощь оказать не может, она обращалась как это было и в Индии к врачу-мужчине. Деятельность греческих акушерок была довольно разнообразной: они не только оказывали помощь при родах и в послеродовом периоде, по занимались и прерыванием беременности. В Древней Греции прерывание беременности в ранних сроках не преследовалось. Эту операцию допускал знаменитый древнегреческий философ и естествоиспытатель Аристотель, считая, что в ранний период развития беременности плод не имеет еще сознания.




 Исправление неправильного положения плода путем поворота, это великое достижение древности, было забыто или не использовалось большинством врачей.
По законам средневековой христианской церкви следовало, что смерть роженицы происходит по воле бога: уничтожение внутриутробного плода, обладающего «божественной душой», рассматривалось как обычное убийство. В итоге церковь сильно препятствовала развитию акушерства, что приводило в конечном счете к высокой смертности в родах.
Возрождение.
16 век
Только в XVI веке,в 1513 году появилось первое, снабжённое рисунками, руководство по акушерству Евхария Ресслина «Цветник беременных женщин и акушерок»; за ним последовали сочинения Якова Руфа в Цюрихе, Вальтера Рейфа в Страсбурге.
При всём том вследствие неполноты научных наблюдений врачей и успехи науки ограничивались только оперативной частью, так как мужчины-врачи приглашались к родам только в очень трудных случаях.
Большое значение для развития акушерства и гинекологии имела деятельность Везалия, Фарлопия, Амбруаза Паре, который вернул акушерству забытую операцию поворота плода на ножку, ввел в широкую практику гинекологические зеркала и организовал при парижском госпитале Hotel-Dieu первое родовспомогательное отделение и первую в Европе акушерскую школу.
Исправление неправильного положения плода путем поворота, это великое достижение древности, было забыто или не использовалось большинством врачей.
По законам средневековой христианской церкви следовало, что смерть роженицы происходит по воле бога: уничтожение внутриутробного плода, обладающего «божественной душой», рассматривалось как обычное убийство. В итоге церковь сильно препятствовала развитию акушерства, что приводило в конечном счете к высокой смертности в родах.
Возрождение.
16 век
Только в XVI веке,в 1513 году появилось первое, снабжённое рисунками, руководство по акушерству Евхария Ресслина «Цветник беременных женщин и акушерок»; за ним последовали сочинения Якова Руфа в Цюрихе, Вальтера Рейфа в Страсбурге.
При всём том вследствие неполноты научных наблюдений врачей и успехи науки ограничивались только оперативной частью, так как мужчины-врачи приглашались к родам только в очень трудных случаях.
Большое значение для развития акушерства и гинекологии имела деятельность Везалия, Фарлопия, Амбруаза Паре, который вернул акушерству забытую операцию поворота плода на ножку, ввел в широкую практику гинекологические зеркала и организовал при парижском госпитале Hotel-Dieu первое родовспомогательное отделение и первую в Европе акушерскую школу.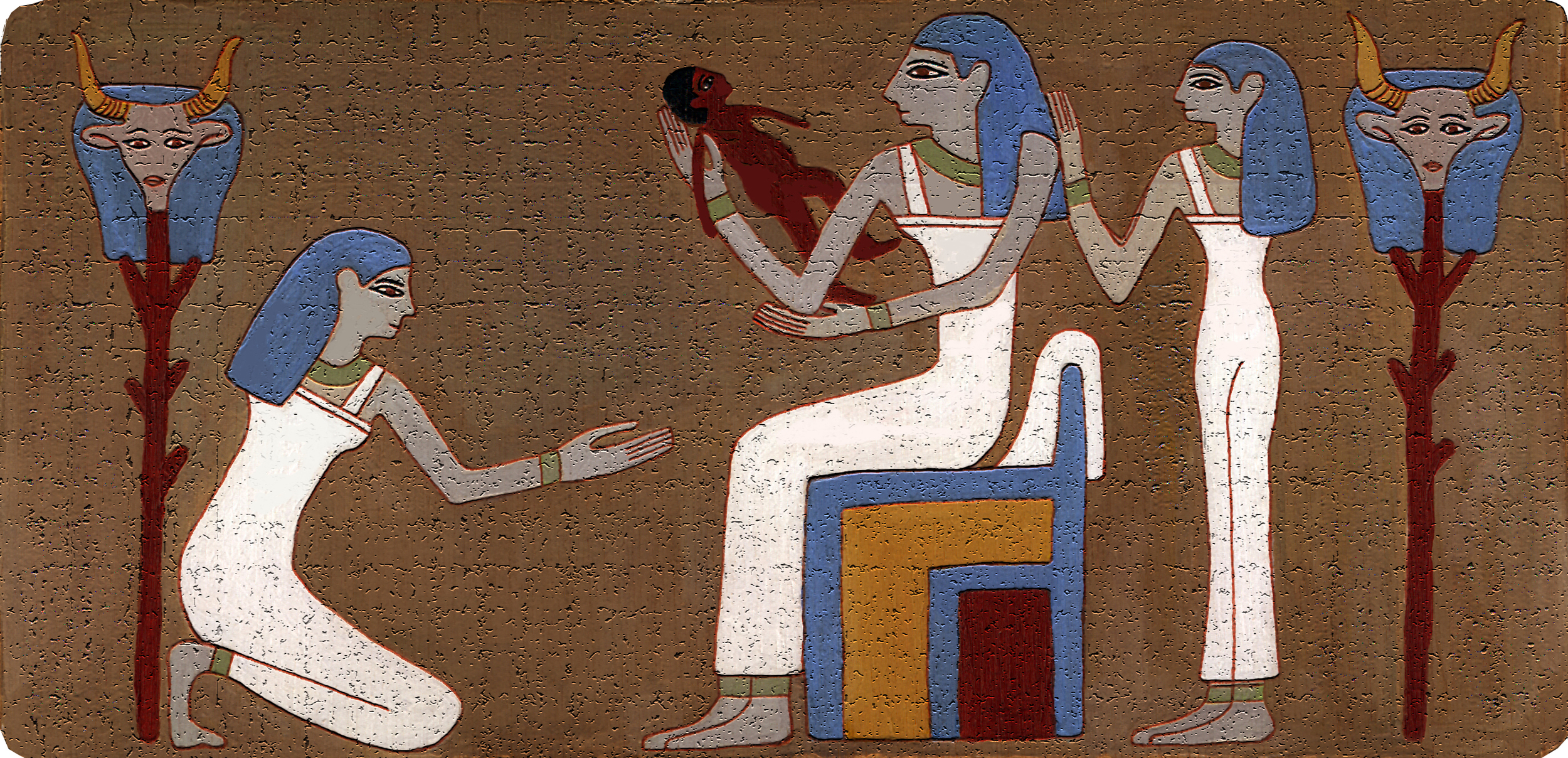 В нее принимались только женщины; обучение длилось 3 месяца, из них 6 недель отводилось на практические занятия.
В этот период стала бурно развиваться анатомия. Это привело к большому числу открытий также в области гинекологии.
Развивающиеся наука и медицина этого периода позволяли проводить достаточно сложные полостные и гинекологические операции.
Разрабатывались новые диагностические методы, позволявшие определить правильность, срок течения родов, а также состояние плода. Изучались анатомические понятия, что позволяло в последующем более или менее точно предсказать течение родов, и соответственно быть готовым ко всем неприятностям.
16 век-подушки
С развитием хирургии продвинулось вперёд и акушерство, особенно во Франции, где врачи всё более и более завоёвывали акушерскую практику. Предрассудки против акушерства, по крайней мере в высших слоях, рассеялись под влиянием приглашения Людовиком XIV знаменитого хирурга Клемента из Арея для оказания акушерской помощи госпоже Лавальер и вскоре назначенного первым акушером королевского двора.
В нее принимались только женщины; обучение длилось 3 месяца, из них 6 недель отводилось на практические занятия.
В этот период стала бурно развиваться анатомия. Это привело к большому числу открытий также в области гинекологии.
Развивающиеся наука и медицина этого периода позволяли проводить достаточно сложные полостные и гинекологические операции.
Разрабатывались новые диагностические методы, позволявшие определить правильность, срок течения родов, а также состояние плода. Изучались анатомические понятия, что позволяло в последующем более или менее точно предсказать течение родов, и соответственно быть готовым ко всем неприятностям.
16 век-подушки
С развитием хирургии продвинулось вперёд и акушерство, особенно во Франции, где врачи всё более и более завоёвывали акушерскую практику. Предрассудки против акушерства, по крайней мере в высших слоях, рассеялись под влиянием приглашения Людовиком XIV знаменитого хирурга Клемента из Арея для оказания акушерской помощи госпоже Лавальер и вскоре назначенного первым акушером королевского двора.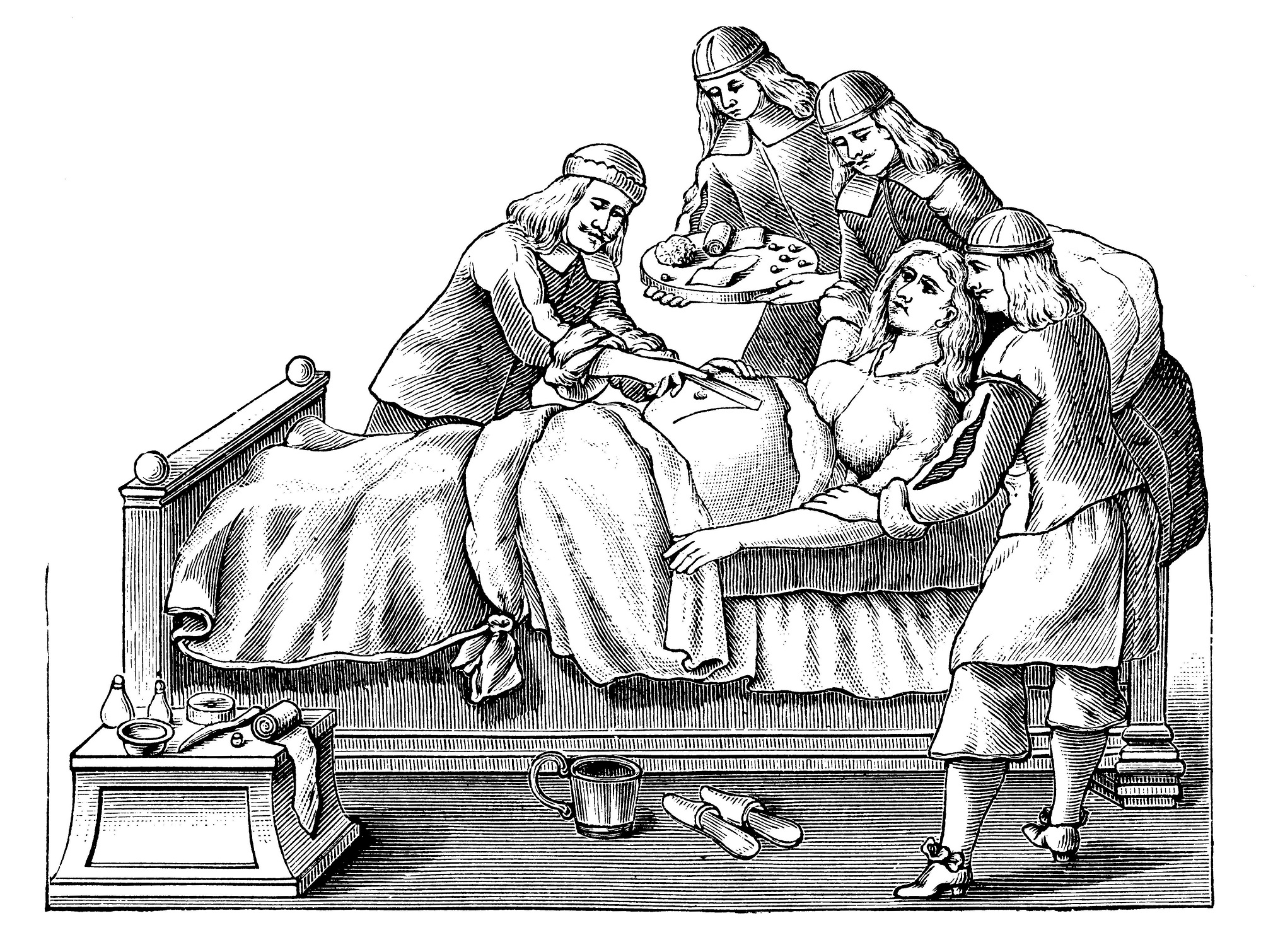 Такое отличие приободрило французских врачей к дальнейшему развитию акушерства.
17 век
Становление акушерства как самостоятельной клинической дисциплины началось во Франции на рубеже XVII-XVIII вв. В значительной степени этому способствовала организация акушерских клиник. Первая из них была открыта в Париже (XVII в.) в госпитале Hotel-Dieu. Здесь сформировалась первая школа французских акушеров, предложивших несколько новых акушерских операций и инструментов.
XVIII век явился периодом становления акушерства в Англии, Голландии, Германии, Франции, России и других странах. Так, в 1729 г. в Страсбурге был открыт первый в Европе родильный дом-клиника. В 1751 г. в Геттингене была организована первая университетская акушерская клиника, где обучали студентов.
В Париже существовало только одно училище для акушерок. В 1728 году открыто родовспомогательное заведение в Страсбурге,
В Англии первое родовспомогательное заведение было открыто в 1765 году.
Такое отличие приободрило французских врачей к дальнейшему развитию акушерства.
17 век
Становление акушерства как самостоятельной клинической дисциплины началось во Франции на рубеже XVII-XVIII вв. В значительной степени этому способствовала организация акушерских клиник. Первая из них была открыта в Париже (XVII в.) в госпитале Hotel-Dieu. Здесь сформировалась первая школа французских акушеров, предложивших несколько новых акушерских операций и инструментов.
XVIII век явился периодом становления акушерства в Англии, Голландии, Германии, Франции, России и других странах. Так, в 1729 г. в Страсбурге был открыт первый в Европе родильный дом-клиника. В 1751 г. в Геттингене была организована первая университетская акушерская клиника, где обучали студентов.
В Париже существовало только одно училище для акушерок. В 1728 году открыто родовспомогательное заведение в Страсбурге,
В Англии первое родовспомогательное заведение было открыто в 1765 году. В Германии первое училище для акушерок учредил в Берлине в 1751 г. Фридрих Великий; в том же году было открыто второе.
По картинкам видно, что рожали как лежа, так и сидя- на краешке кровати, на подушках или на специальных стульях-креслах. Иногда просто спускались на корточки.
Картинки разложены по времени, и по ним видно, что где не было кресел- рожали лежа.
С развитием медицины расширялся арсенал действий акушеров, что требовало горизонтального положения.
По картинкам видно, что рожали как лежа, так и сидя- на краешке кровати, на подушках или на специальных стульях-креслах. Иногда просто спускались на корточки.
Картинки разложены по времени, и по ним видно, что где не было кресел- рожали лежа.
С развитием медицины расширялся арсенал действий акушеров, что требовало горизонтального положения.
Родильные кресла
Родильные стулья использовались с древнейших времен до 1800-х годов.Они часто передавались в семьях по наследству.
В Германии первое училище для акушерок учредил в Берлине в 1751 г. Фридрих Великий; в том же году было открыто второе.
По картинкам видно, что рожали как лежа, так и сидя- на краешке кровати, на подушках или на специальных стульях-креслах. Иногда просто спускались на корточки.
Картинки разложены по времени, и по ним видно, что где не было кресел- рожали лежа.
С развитием медицины расширялся арсенал действий акушеров, что требовало горизонтального положения.
По картинкам видно, что рожали как лежа, так и сидя- на краешке кровати, на подушках или на специальных стульях-креслах. Иногда просто спускались на корточки.
Картинки разложены по времени, и по ним видно, что где не было кресел- рожали лежа.
С развитием медицины расширялся арсенал действий акушеров, что требовало горизонтального положения.
Родильные кресла
Родильные стулья использовались с древнейших времен до 1800-х годов.Они часто передавались в семьях по наследству. Родовые кресла были на пике своей популярности к концу 1700-х годов, использовались и женщинами, и мужчинами- акушерами.
Спинка в родильном кресле, позволяет матери сидеть прямо во время родов. Тем не менее, оно имеет низкий центр тяжести, поэтому мать может также сесть на корточки и по-прежнему опираться на стул.
Многие стулья были семейными реликвиями, которые передаются из поколения в поколение по сегодняшний день. Они все еще используются в некоторых странах.
Некоторые стулья имели довольно сложные конструкции и подставки для ног для комфорта матери. Некоторые также могут быть сложены, или преобразованы в кровать для операционной.
Этот стул был использован для примерно 2000 родов -был распоряжении семьи акушерок на протяжении трех поколений. Образ Христа на спинке стула- возможно, был знаком защиты для женщин и их детей, поскольку роды были рискованным событием.
Сиденья имеет форму, чтобы разрешить доступ во время родов, также может быть сложено преобразовано в кровать или операционный стол.
Родовые кресла были на пике своей популярности к концу 1700-х годов, использовались и женщинами, и мужчинами- акушерами.
Спинка в родильном кресле, позволяет матери сидеть прямо во время родов. Тем не менее, оно имеет низкий центр тяжести, поэтому мать может также сесть на корточки и по-прежнему опираться на стул.
Многие стулья были семейными реликвиями, которые передаются из поколения в поколение по сегодняшний день. Они все еще используются в некоторых странах.
Некоторые стулья имели довольно сложные конструкции и подставки для ног для комфорта матери. Некоторые также могут быть сложены, или преобразованы в кровать для операционной.
Этот стул был использован для примерно 2000 родов -был распоряжении семьи акушерок на протяжении трех поколений. Образ Христа на спинке стула- возможно, был знаком защиты для женщин и их детей, поскольку роды были рискованным событием.
Сиденья имеет форму, чтобы разрешить доступ во время родов, также может быть сложено преобразовано в кровать или операционный стол. После родов
17 век
18-век
19 век
Диагностика по моче
Россия
В России наука акушерства получила своё начало и развитие значительно позже, чем в других европейских странах. Первый акушер, о котором упоминается в летописях, был англичанин Якоб (при Иоанне Грозном), славившийся как «умеющий очень искусно лечить женские болезни».
Хотя Петром Великим открыты были школы «для медической и хирургической практики», но так как имелись в виду исключительно потребности армии и флота, акушерству в них не обучали. Только в 1754 году устроены были акушерские школы в Петербурге и Москве, преподавать в которых были приглашены профессора с помощниками, в Петербурге Линдеман, в Москве Эразмус, которому принадлежит первое акушерское сочинение на русском языке — «Наставление, как женщине в беременности, в родах и после родов себя содержать надлежит». Означенное руководство было составлено по Горну, оригинал которого был издан в 1697 году, так что в середине прошлого столетия, когда уже Смелли, Левре и Редерер преобразовали акушерство, русские врачи черпали свои знания из давно устаревшей книги конца XVI столетия.
После родов
17 век
18-век
19 век
Диагностика по моче
Россия
В России наука акушерства получила своё начало и развитие значительно позже, чем в других европейских странах. Первый акушер, о котором упоминается в летописях, был англичанин Якоб (при Иоанне Грозном), славившийся как «умеющий очень искусно лечить женские болезни».
Хотя Петром Великим открыты были школы «для медической и хирургической практики», но так как имелись в виду исключительно потребности армии и флота, акушерству в них не обучали. Только в 1754 году устроены были акушерские школы в Петербурге и Москве, преподавать в которых были приглашены профессора с помощниками, в Петербурге Линдеман, в Москве Эразмус, которому принадлежит первое акушерское сочинение на русском языке — «Наставление, как женщине в беременности, в родах и после родов себя содержать надлежит». Означенное руководство было составлено по Горну, оригинал которого был издан в 1697 году, так что в середине прошлого столетия, когда уже Смелли, Левре и Редерер преобразовали акушерство, русские врачи черпали свои знания из давно устаревшей книги конца XVI столетия. ——————————————
В 1757 г. в Москве и Петербурге были созданы «бабичьи школы», которые готовили «присяжных бабок» (образованных повивальных бабок, или акушерок). Преподавали в них первоначально иностранцы: один доктор (профессор бабичьего дела) и один лекарь (акушер). В первые годы обучение было только теоретическим. Затем, после открытия в России повивальных (родильных) отделений на 20 коек при Московском и Петербургском воспитательных домах, стал преподаваться и практический курс. Сначала обучение в бабичьих школах было малоэффективным. Имелись существенные трудности при наборе учениц: так, в 1757 г. в Петербурге были зарегистрированы 11, а в Москве — 4 повивальные бабки, — именно они составляли весьма ограниченный резерв набора учащихся. В результате за первые 20 лет Московская «бабичья школа» подготовила всего 35 повивальных бабок ,из которых пять были из «прирожденных россиян», а остальные — иностранки.
осмотр :
——————————————
В 1757 г. в Москве и Петербурге были созданы «бабичьи школы», которые готовили «присяжных бабок» (образованных повивальных бабок, или акушерок). Преподавали в них первоначально иностранцы: один доктор (профессор бабичьего дела) и один лекарь (акушер). В первые годы обучение было только теоретическим. Затем, после открытия в России повивальных (родильных) отделений на 20 коек при Московском и Петербургском воспитательных домах, стал преподаваться и практический курс. Сначала обучение в бабичьих школах было малоэффективным. Имелись существенные трудности при наборе учениц: так, в 1757 г. в Петербурге были зарегистрированы 11, а в Москве — 4 повивальные бабки, — именно они составляли весьма ограниченный резерв набора учащихся. В результате за первые 20 лет Московская «бабичья школа» подготовила всего 35 повивальных бабок ,из которых пять были из «прирожденных россиян», а остальные — иностранки.
осмотр :
Как склоняется слово «род» — Российская газета
Род роду рознь.
Это и так понятно, но еще более очевидно становится, когда заглянешь в словари. Во-первых, есть «рОды», которые всегда во множественном числе, — это процесс рождения ребенка. Единственного числа здесь нет вообще, да и с ударением все ясно. Дальше — еще интереснее. В словарях сейчас есть три совершенно одинаковых на вид, но разных по значению слова «род».
Давайте начнем с «рода», который означает ряд поколений или, как пишет словарь, первобытную общественную организацию. Во множественном числе они — «родЫ». Есть и другой «род» — тот, которым мы называем единицу классификации. Род и вид, рОды и виды. В русском языке три рОда — мужской, женский, средний. Род, рОды, по родАм.
А бывают еще и родА войск или оружия. Потому что они не «рОды» и не «родЫ», а родА. Именно родА войск, родА оружия. «Сколько родОв оружия вы знаете? Немного? Это, поверьте, к лучшему!»
Вот так: рОды, родЫ, родА. Похоже на карнавал с переодеваниями. Наша задача — не перепутать маски.
Радушный и радужный
Пишутся и звучат эти слова похоже. Но они, конечно же, очень разные. «РадУшный» происходит от слова «радушие» (открытое, сердечное расположение к людям, к гостям).
Но они, конечно же, очень разные. «РадУшный» происходит от слова «радушие» (открытое, сердечное расположение к людям, к гостям).
Интересно, что «радушие» есть только в русском и болгарском языках, в других славянских языках это слово отсутствует. Зато уж в русском замечено с начала XVIII века! Предполагают, что возникло оно из слова «радодушие» — так же, как в свое время «курносый» из «корноносый».
«Радужный» — это от «радуги», разноцветной полосы на небосводе. Однако в древнерусском языке, как выясняется, не было слова «радуга», было просто «дуга». Языковеды не исключают, что это слово тоже образовалось из двух: «радо-дуга», так же как «радо-душие».
Вот видите, начинали мы с того, что «радушный» и «радужный» — слова совсем разные, но этимология коварна: стоит в нее углубиться, и все слова оказываются в какой-то степени родственниками.
Однако это было в далеком прошлом, сейчас их все же лучше не путать.
Городская клиническая больница имени Е.О.Мухина
Рефлексотерапия, или акупунктура, известна человечеству с давних времен. Традиционно родиной этого метода лечения считаются древний Китай и Корея. Именно в древних китайских трактатах (6-5 вв. до н.э.) впервые упоминается иглоукалывание как метод лечения многих заболеваний. Однако некоторые археологические находки позволяют предположить, что акупунктура возникла гораздо раньше. Найденные при раскопках костяные, каменные и бамбуковые иглы датируются периодом более чем 5 тыс. лет до н.э. Согласно китайской философии, тело человека пронизывают невидимые энергетические каналы, распределяющие живительную энергию по всему человеческому телу. Именно с помощью иглоукалывания в точках акупунктуры можно воздействовать на эти каналы, восстанавливая нарушенный энергетический баланс, тем самым восстанавливая и здоровье
Традиционно родиной этого метода лечения считаются древний Китай и Корея. Именно в древних китайских трактатах (6-5 вв. до н.э.) впервые упоминается иглоукалывание как метод лечения многих заболеваний. Однако некоторые археологические находки позволяют предположить, что акупунктура возникла гораздо раньше. Найденные при раскопках костяные, каменные и бамбуковые иглы датируются периодом более чем 5 тыс. лет до н.э. Согласно китайской философии, тело человека пронизывают невидимые энергетические каналы, распределяющие живительную энергию по всему человеческому телу. Именно с помощью иглоукалывания в точках акупунктуры можно воздействовать на эти каналы, восстанавливая нарушенный энергетический баланс, тем самым восстанавливая и здоровье
Лечение иглоукалыванием кажется простым, но это только на наш непросвещенный взгляд, на самом же деле этот метод требует не только огромных познаний в строении человеческого тела, но и ювелирного владения техникой данного метода. Иглорефлексотерапия официально признана как терапевтический метод Всемирной организацией здравоохранения (ВОЗ). Список заболеваний, при которых она рекомендуется, согласно клиническим испытаниям состоит из 28 пунктов. Ученые тоже отмечают, что при воздействии иглы на особые точки на коже происходит мгновенный рефлекторный ответ соответствующих им внутренних органов. Его силой и характером можно управлять. Так и происходит лечение различных заболеваний при помощи иглоукалывания.
Список заболеваний, при которых она рекомендуется, согласно клиническим испытаниям состоит из 28 пунктов. Ученые тоже отмечают, что при воздействии иглы на особые точки на коже происходит мгновенный рефлекторный ответ соответствующих им внутренних органов. Его силой и характером можно управлять. Так и происходит лечение различных заболеваний при помощи иглоукалывания.
С помощью иглоукалывания опытному специалисту удается без медикаментов воздействовать на все основные органы и системы человеческого организма. Причем метод иглоукалывания можно сочетать с другими видами лечения, дополняя и усиливая их. К тому же этот метод имеет очень ограниченные противопоказания и не вызывает никаких аллергических реакции.
Метод иглоукалывания улучшает обменные процессы в организме, стимулирует выработку собственных гормонов, усиливает защитные реакции организма. Применяется этот метод довольно широко, вот примерный перечень заболеваний, при которых возможно добиться положительных результатов с помощью игл.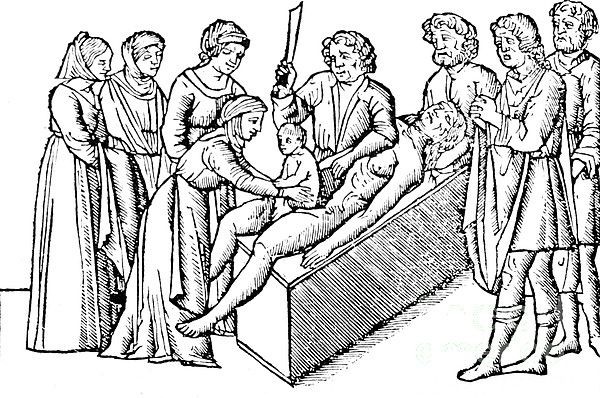 нервные окончания связаны не только с каким-то определенным органом тела, но, прежде всего, с нервной системой и основным ее органом – головным мозгом. Именно головной мозг, в процессе воздействия игл, тем или иным образом реагирует на такое раздражение. И именно это делает возможность иглоукалыванию приносить существенную пользу здоровью в тех случаях, когда ничего другое помочь не может.
нервные окончания связаны не только с каким-то определенным органом тела, но, прежде всего, с нервной системой и основным ее органом – головным мозгом. Именно головной мозг, в процессе воздействия игл, тем или иным образом реагирует на такое раздражение. И именно это делает возможность иглоукалыванию приносить существенную пользу здоровью в тех случаях, когда ничего другое помочь не может.
Как проводится сеанс иглорефлексотерапии
Обычно на один сеанс ставят 8-10 иголок, которые находятся в теле пациента 20-40 минут. К концу процедуры некоторые иголки сами выходят из тканей либо врач извлекает их безболезненно для пациента. Кратность и длительность лечения определяются индивидуально, но в любом случае следует быть готовым к курсу из 10-12 процедур, которые проводятся ежедневно. Для лечения впервые возникших патологий в комплексной терапии достаточно 1 курса иглотерапии, хронические же патологии требуют курсового лечения иголками 1 раз в квартал. Некоторые пациенты отмечают облегчение уже после первого сеанса иглорефлексотерапии, другим же требуется несколько недель после завершения сеанса, чтобы организм ответил положительным эффектом. В любом случае, эффект от процедуры – индивидуальный показатель, который зависит от состояния организма.
В любом случае, эффект от процедуры – индивидуальный показатель, который зависит от состояния организма.
Что ощущает пациент при постановке иголок? В целом процедура переносится хорошо и не причиняет дискомфорта, а тем более выраженных болей. Ощущения при постановке иголок индивидуальны и зависят от индивидуального порога чувствительности. Иглы имеют закругленный край, поэтому они не прокалывают кожу, а раздвигают ее. Во время процедуры человек ощущает пульсацию и небольшое давление в акупунктурных точках. Возможно чувство незначительного покалывания, ощущения зуда и тепла.
- заболевания центральной и периферической нервной системы
- заболевания позвоночника, сопровождаемые болью в различных его отделах
- боли неврологического характера, такие как межреберная невралгия и невралгия лицевого нерва
- головные боли различного происхождения, в том числе мигрень
- мышечные боли
- воспалительные заболевания суставов
- нарушение обмена веществ, в том числе сахарный диабет и ожирение
- снижение иммунитета
- депрессия
- хроническая усталость
- нарушение сна
- заболевания желудка и кишечника, к том числе колиты, гастриты, язва желудка и двенадцатиперстной кишки
- заболевания печени
- заболевания мочеполовой системы
- расстройства сексуальной сферы
- бронхиальная астма
- заболевания легких
- гинекологические заболевания
- частые простуды, болезни горла
- аллергические реакции
- алкогольная зависимость
Древние жители Руси
История земель, которые сегодня считаются исконно российскими, началась задолго до появления у восточных славян государства. Русскую равнину обжили еще 25 тысяч лет назад — под Владимиром нашли стоянку древнего человека этого периода. На территории нашей страны жили предки балтов и германцев, а первые «москвичи» были из финно-угорских племен. Портал «Культура.РФ» собрал 7 интересных фактов о жителях Центральной России до появления здесь славянских народов.
Русскую равнину обжили еще 25 тысяч лет назад — под Владимиром нашли стоянку древнего человека этого периода. На территории нашей страны жили предки балтов и германцев, а первые «москвичи» были из финно-угорских племен. Портал «Культура.РФ» собрал 7 интересных фактов о жителях Центральной России до появления здесь славянских народов.
Первые стоянки Русской равнины
Жители стоянки Сунгирь изготавливают украшения. Изображение: arheo-news.ruСчитается, что люди обжили Русскую равнину в верхнем палеолите. К этому периоду относится стоянка древнего человека — Сунгирь под Владимиром. Возраст стоянки — около 25 тысяч лет. Это было сезонное охотничье стойбище, которым пользовались, как полагают ученые, на протяжении двух-трех тысяч лет. Сегодня этот памятник находится под охраной ЮНЕСКО.
В поселении археологи обнаружили захоронение двух мальчиков — 12 и 14 лет. Здесь также нашли кость взрослого человека, наполненную охрой. Исследователи выяснили, что кость принадлежала прапрадедушке похороненных подростков и имела особое значение в погребении: мальчиков, скорее всего, принесли в жертву в честь культа плодородия.
Еще 5 археологических памятников России
В могилах были копья и дротики из бивня мамонта, а также диски, символизирующие солнце. Одежда детей была вышита бусинами из бивня мамонта — ученые нашли их около 10 тысяч штук. Наряды напоминали костюмы нынешних северных народов, а после реконструкции их внешнего облика стало ясно, что сунгирские люди могли быть предками современных североевропейцев.
Европейцы-кочевники
Каменные сверленые топоры фатьяновской культуры. Фотография: tm-arh.ruВо III–II тысячелетии до н. э. на территории Центральной России жили высокие люди европейского типа с широкими лицами. Они относились к общности, из которой позднее выделились балты, германцы и славяне. Эту археологическую культуру назвали фатьяновской — по могильнику, который открыл археолог Алексей Уваров в 1873 году. Ученый обнаружил его у деревни Фатьяново (сегодня — Ярославский район). Второе название — «культура боевых топоров» — возникло благодаря обычаю этих людей класть в мужские могилы топоры, выточенные из камня. Хоронили они, кстати, не только людей, но и животных — в основном медведей и собак. Их фатьяновцы почитали как предков своего рода.
Хоронили они, кстати, не только людей, но и животных — в основном медведей и собак. Их фатьяновцы почитали как предков своего рода.
Фатьяновцы кочевали, устраивая легкие жилища, разводили свиней, овец и коз, изготавливали костяные и каменные мотыги. Имущество они перевозили на повозках и телегах.
Ученые нашли следы кочевников в Ивановской и Ярославской, Тверской и Костромской, Нижегородской и Владимирской, Рязанской и Тульской областях, а также в предгорьях Урала. Со временем фатьяновцев стали теснить наступавшие с востока племена — часть народа отступила на запад, а другая часть смешалась с захватчиками.
Первые москвичи
Предметы дьяковской культуры и древнерусского времени. Фотография: nasledie.dubna.ruС VIII–VII веков до н. э. земли от Вологды до Смоленска населяла дьяковская археологическая культура. Только в границах современной Москвы обнаружено 10 дьяковских городищ — все они были построены на высоких мысах при слиянии рек. Именно так возникло древнейшее поселение на месте Московского Кремля. Известно, что дьяковцы относились к финно-угорским племенам. Именно от их потомков — племен меря и весь — нам достались многие названия рек: Яхрома, Кашира, Вологда, Вычегда.
Известно, что дьяковцы относились к финно-угорским племенам. Именно от их потомков — племен меря и весь — нам достались многие названия рек: Яхрома, Кашира, Вологда, Вычегда.
Дьяковцы вели оседлый образ жизни — в каждом городище проживало от 50 до 200 человек. Примерно с IV века до н. э. широко распространилось железо и выросло благосостояние родов, а поэтому участились и грабительские набеги. Дьяковцы стали укреплять свои поселения частоколами, земляными валами и рвами. Основным занятием их было скотоводство: они разводили лошадей. Причем в основном на еду, как тягловую силу коней практически не использовали. Также население охотилось: на лосей и оленей, медведей и кабанов. Шкурки бобров, лис, куниц и выдр использовали как ходовую валюту во время обмена с другими племенами.
Мертвых дьяковцы сжигали и хоронили в «домиках мертвых». Такие захоронения нашли в Березняках на реке Волге (сегодня Ярославская область), недалеко от Саввино-Сторожевского монастыря в Московской области. Одна из гипотез говорит, что сказочная избушка на курьих ножках Бабы-яги — это найденные в лесу «домики» дьяковцев.
Одна из гипотез говорит, что сказочная избушка на курьих ножках Бабы-яги — это найденные в лесу «домики» дьяковцев.
Прародина славян
Археологи и языковеды установили, что славяне отделились от древнеевропейской общины в 400 году до н. э. К тому времени уже существовали кельты и италики, германцы и западные балты, венеты и иллирийцы. По одной версии, прародиной славян была долина между реками Вислой и Одрой (Одером) на территории современной Польши. Другие ученые предполагают, что славяне изначально селились между Западным Бугом и средним течением Днепра — сегодня это территория на пересечении Польши, Украины и Беларуси. Долгое время считалось, что предки славянских народов пришли с Дуная — эта теория была основана на сведениях из Повести временных лет. Сегодня ученые признали ее ненаучной.
А здесь читайте о божествах и ритуалах древних славян
Североевропейское происхождение славян неожиданно подтвердил древнеанглийский язык. В нем было много славизмов — заселившие в IV–V веке Британские острова англы, саксы и юты прежде жили на датском полуострове Ютландия и нижней Эльбе. Их соседями были славяне.
Их соседями были славяне.
«Великая славянская миграция»
Сергей Иванов. Жилье восточных славян. Иллюстрация к сборнику «Картины по русской истории». Издание Иосифа Кнебеля. 1909
В IV веке на земли готов и римлян вторглись гунны — азиатские кочевники, занявшие Юго-Восточную и Центральную Европу. Спасаясь от них, европейцы массово бежали на запад, тесня другие племена. Так на протяжении почти трех веков проходило Великое переселение народов. В учебниках истории миграцию славянских народов объясняют именно этим процессом, но археологи подчеркивают: славяне стали расселяться на юг и восток еще до гуннов, в начале новой эры. В VI веке они уже составляли основную часть населения Аварского каганата — государства, основанного в Центральной Европе аварами.
Настоящая «великая славянская миграция» была спровоцирована похолоданием, начавшимся в конце IV века. Самым холодным за прошедшие 2000 лет был V век. В это время поднялся уровень воды в Северном и Балтийском морях, реки затопили прибрежные поселения. Из-за затопленных полей и разрастания болот люди стали массово покидать Висло-Одерский регион — свою исконную территорию. К VII–VIII векам они пересекли границу современной России.
Подмосковные прибалты
В IX веке к моменту образования Древнерусского государства на территории нынешней Центральной России было смешанное население. Коренными на тот момент были финно-угры и балты, иноземными — славяне и варяги. В Повести временных лет летописец перечислял племена, «дающие дань Руси»: весь, меря, мурома, черемисы, мордва, чудь, пермь, печера, ямь, литва, зимиголы, корсь, нарова и ливы.
На границе Московской, Калужской и Смоленской областей жило племя голядь, которое окончательно ассимилировалось лишь в XIV веке. Вероятно, сами себя представители этого народа называли галиндами, а пришли они из прусской области Галиндии. Говорили они на языке, родственном литовскому и латышскому. Переселившись на Оку во II веке, галинды быстро смешались с обитавшими здесь восточными балтами. На память об этом народе нам остались балтские названия подмосковных рек: Ока, Дубна, Протва и Истра. По одной из версий, и слово «Москва» имеет балтийский корень.
На память об этом народе нам остались балтские названия подмосковных рек: Ока, Дубна, Протва и Истра. По одной из версий, и слово «Москва» имеет балтийский корень.
Какие славянские племена жили в России
Восточные славяне. Фотография: sokrovennik.ruВ Повести временных лет автор упомянул 15 славянских племенных союзов — на территории современной России жили три: словене, кривичи и вятичи. Словене основали Великий Новгород, Ладогу, Белоозеро, Старую Руссу и Псков. В 862 году в Новгород пригласили варяга Рюрика — этот момент связывают с возникновением российской государственности.
«Земля наша велика и обильна, а наряда в ней нет. Приходите княжить и воладети нами».
Домовые и лешие, упыри и русалки — во что верили наши предки
Кривичи расселились от Днепра и Западной Двины до верхней Волги. Их главными городами были Смоленск, Изборск и Полоцк. Преемник Рюрика Вещий Олег, перенесший столицу в Киев в 882 году, покорил Смоленск, но кривичи сопротивлялись «центральной власти» еще почти 100 лет.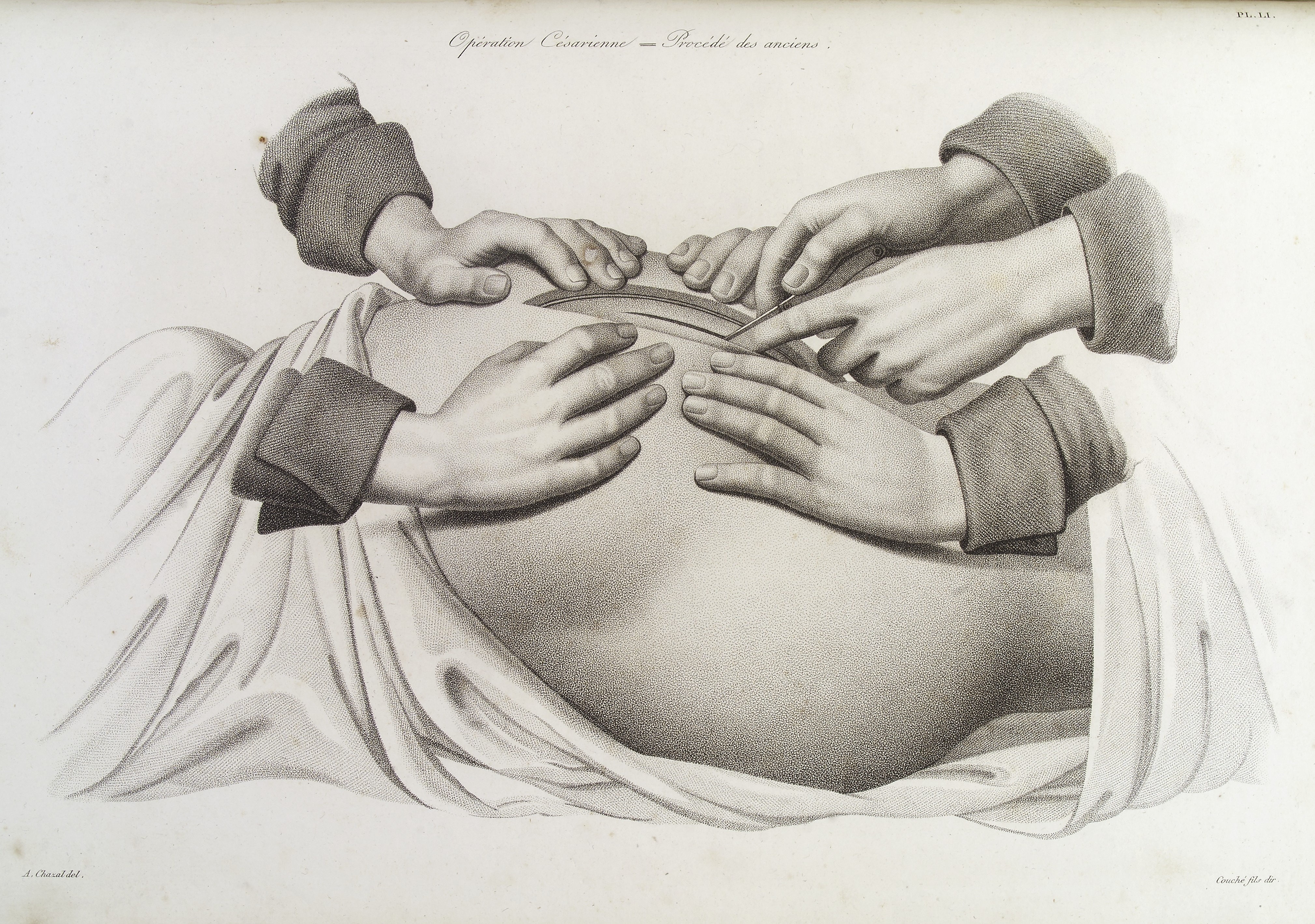 Главным городом вятичей, живших по Оке и верховью Дона, стал Корьдно недалеко от современной Калуги. На момент образования Древнерусского государства они платили дань хазарам и жили обособленно. Окончательно присоединить земли вятичей к Древней Руси удалось лишь в XI веке.
Главным городом вятичей, живших по Оке и верховью Дона, стал Корьдно недалеко от современной Калуги. На момент образования Древнерусского государства они платили дань хазарам и жили обособленно. Окончательно присоединить земли вятичей к Древней Руси удалось лишь в XI веке.
Автор: Екатерина Гудкова
give birth — Translation into Russian — examples English
Suggestions: give birth to women who give birthThese examples may contain rude words based on your search.
These examples may contain colloquial words based on your search.
You can give birth for our country.
Вы можете рожать для нашей страны.What, give birth in prison?
She was 16 and about to give birth.
Number of young women due to give birth nine months later.
Количество молодых женщин, собирающихся родить через девять месяцев.Others give birth after wearing the wedding dress,
Другие рожают после того, как надели свадебное платье, а у меня всё наоборот.Mr Amos has a young wife about to give birth.
У мистера Амоса молодая жена, которая должна вот-вот родит.
I once saw a zebra named Gavin give birth at the zoo, and I cried hysterically.
She’s saying that she wants to give birth at home.
Она сказала, что будет рожать дома.Indians give birth alone and standing.
Древние индианки рожали в одиночку и стоя.The doctor told Nili she could give birth any day now.
Доктор сказал Нили, что она может родить в любую минуту.
I actually do know a technique that would allow you to give birth naturally.
And watching that girl give birth at prom is what made me want to be an obstetrician.
И увидев, как та девушка родила на выпускном, я захотела стать акушеркой.You are the first woman to give birth with the help of my fertility clinic.
Ты первая женщина, которая родит с помощью моей клиники планирования семьи.Maybe for her new performance piece, she’ll give birth on-stage.
Может в своем новом спектакле она будет рожать на сцене.
I can’t give birth without my mother here.
Я не могу рожать без моей матери рядом.It means you’re ready to give birth.
I give birth in rain at side of road.
Я родить под дождем у края дороги.I think she’ll give birth tonight.
Я думаю, она родит сегодня ночью.We can only conceive and give birth in the water.
Мы можем зачать и родить только в воде.
We need a guy who can teach them, how to give birth, only to male children.
Им нужен поводырь, который может научить их рожать только детей мужского пола. ДИНАСТИИГЕНЕАЛОГИЯГЕОГРАФИЯЛИТЕРАТУРАРодственные проекты:ХРОНОСФОРУМ ХРОНОСАПРАВИТЕЛИ МИРАВОЙНА 1812 ГОДАОТ НИКОЛАЯ ДО НИКОЛАЯПЕРВАЯ МИРОВАЯРЕПРЕССИРОВАННОЕ ПОКОЛЕНИЕДОКУМЕНТЫ XX ВЕКАПрочее: | Дворянские роды РоссииАндогские — русский княжеский род, Рюриковичи, составляли ветвь князей Белозерских. Название приняли от Андогской волости,
находившейся на территории современной
Новгородской области, в 80 километрах от
Белозерска. Основателем рода был князь Михаил,
сын белозерского князя Андрея Юрьевича. Первым
из известных андогских князей становится
Александр, сын Михаила, которому верейский князь Михаил Андреевич
продал в 1486 году некоторые земли не Белозере, а
после него — племянник Григорий Иванович,
который, будучи сыном боярским, в 1495 году среди
прочих сопровождал в Литву дочь Ивана III
Васильевича Елену. Последний из Андогских князей
упомянут под 1565 годом: Иван Васильевич был
поручителем за Л.А. Салтыкова перед царем. Название приняли от Андогской волости,
находившейся на территории современной
Новгородской области, в 80 километрах от
Белозерска. Основателем рода был князь Михаил,
сын белозерского князя Андрея Юрьевича. Первым
из известных андогских князей становится
Александр, сын Михаила, которому верейский князь Михаил Андреевич
продал в 1486 году некоторые земли не Белозере, а
после него — племянник Григорий Иванович,
который, будучи сыном боярским, в 1495 году среди
прочих сопровождал в Литву дочь Ивана III
Васильевича Елену. Последний из Андогских князей
упомянут под 1565 годом: Иван Васильевич был
поручителем за Л.А. Салтыкова перед царем.
Балакиревы — старинный дворянский род.
Бартеневы — дворянский род.
Бахтеяровы-Ростовские — княжеский род
Безобразовы (1). Владимирская ветвь.
Безобразовы (2). Брянская ветвь.
Безобразовы (3). Смоленская ветвь.
Безобразовы (4). Калужская ветвь
Воронцовы — дворянский род (генеалогическая таблица)
Друцкие — княжеский род.
Друцкие-Соколинские — княжеский род, Рюриковичи, ветвь кн. Друцких, происходившая от внука удел. кн. друцк. Ивана Семёновича Бабы — кн. С. Ф. Бабича, имевшего поместье неподалё- ку от г. Друцк под названием «Сокольня », от к-рого весь его род и получил наименование Соколинских.
Енгалычевы
Отяевы — русский дворянский род.
Родоначальником их был Ф.
Офросимовы — русский дворянский род; по древним родословцам, происходил от «мужа честна» Андрея Андреевича Офросимова, выехавшего из Молдав. ки-ва на службу к вел. кн. Василию II Тёмному. 3. Офросимов служил (1597) осадным головой в Черни.
Ошанины — русский дворянский род, происходивший, по сказаниям родословцев, от выходца из Венеции, Стени, принявшего при крещений в Москве в сер. XIV в. имя Федора. Его потомок в 6-м колене, Даниил Ошаня, стал родоначальником Ошаниных. В.Ф. Ошанин служил ясельничим Ивана Грозного.
Погожевы — русский дворянский род,
происходивший, по древним родословцам, от
литовца Василия Воргоса, по прозвищу Погожий,
выехавшего в Москву к вел.
Пожарские — русский княжеский род, ветвь удел. князей стародуб., происходивших от вел. кн. влад. Всеволода Большое Гнездо. Потомок его 7-го сына, Ивана, родоначальник князей стародубч Василий Андреевич, получил в удел с. Погары и стал писаться князем Пожарским. Род Пожарских ничем особым не выделялся. Широкую известиость в России получил лишь один из Пожарских — известный политический деятель и военачальник в период Смутного времени, руководитель 2-го ополчения — кн. Дмитрий Михайлович Пожарский.
Полевы — русский дворянский род,
происходивший от смол. впадет, князей. У смол. кн.
Федора Юрьевича было 2 сыновей — Борис и Остафий. У
Бориса, князем уже не писавшегося, был
единственный сын Александр, по прозвищу Поле,
служивший боярином при литовском дворе.
Приклонские — русский дворянский род, восходящий к середине XV века. Колупай Михайлович Приклонский был послом к казанскому хану Мухаммеду-Эмину (1492 год). Его сын Михаил ездил в Ногацскую орду (1564). Андрей Гаврилович Приклонский (умер в 1577 году) служил наместником в Курмыше.
Проестевы — русский дворянский род, ветвь новгородского рода Софроновских. Их предок Дмитрий Минин был боярином у великого князя московского Василия I Дмитриевича. Его потомок в шестом колене, Давыд Григорьевич Минин-Сафроновский, имел прозвище Проесть и служил Василию III Ивановичу. Матвей Павлович Проестев служил (1580) воеводой в г. Белый.
Прокудины-Горские — русский
дворянский род, происходивший от одного из
воевод, участвовавших в Куликовской битве (1380), —
Петра Горского, выходца из Золотой Орды.
Прончищевы — русский дворянский род, происходивший от Ивана Васильевича Прончище, выехавшего из Польши к великому князю московскому Ивану III Васильевичу и пожалованного (1488) поместьями в Тарусском уезде. Сын его Яков упомянут в поручной грамоте (1563) за удел. воротын. кн. Александра Ивановича.
Протасьевы — русский дворянский род, происходивший от мценского воеводы Григория Протасьевича. Ивану Григорьевичу Раслу и его сыну Конону была пожалована (1455) в кормление волость Луза. Иван Петрович Келарь был в 1589 наместником в Елатьме и Кадоме.
Протопоповы — русский дворянский род,
происходивший, по древним родословцам, от
выехавшего «из немец» некоего Галцеского,
крестившегося под именем Александра.
Толстые — дворянский род происходит от
древней германской фамилии. Их предком был
Индрис, который в середине XIV века покинул
пределы Германии и вместе двумя сыновьями
поселился в Чернигове. Здесь он принял крещение и
получил имя Леонтия. Родоначальником Толстых
стал правнук Индриса, Андрей Харитонович,
который переселился из Чернигова в Москву и
здесь уже от Василия Темного получил прозвище
Толстой, которое впоследствии стало
передаваться и его потомкам. Первые из
представителей этой фамилии были военными. Эта
традиция сохранилась всеми поколениями Толстых,
однако впоследствии многие Толстые прославили
свой род и как видные государственные чиновники,
и как деятели искусства и литературы.
Уваровы — русский дворянский род, происходивший от выехавшего из Золотой Орды к великому князю московскому Василию I Дмитриевичу мурзы Минчака Касаева, в православном крещении получившего имя Семен. У него были дети: Давид, Злоба, Орешка и Увар, от которых пошли фамилии Давыдовых, Злобиных, Орешкиных и Уваровых. Потомок Увара, Дмитрий Сергеевич Уваров, при Иване Грозном пожалован поместьями в Пскове, откуда был переведён в Белев.
Ухтомские — русский княжеский род,
Рюриковичи. Происходили от белозерск. удел.
князей. Правнук вел. кн. киев. Владимира
Святославича, вел. кн. влад. Всеволод Юрьевич
Большое Гнеяуо, посадил своего сына Константина
в Ростове. Внук Константина, кн. Глеб, был на уделе
в Белоозере. У правнука Глеба, кн. Василия
Романовича, был внук Иван, владевший волостью на
берегах р.
Ушатые — русский княжеский род, Рюриковичи. Происходили от ярославских князей. Сын вел. кн. влад. Всеволода Юрьевича Большое Гнездо Константин имел сына Всеволода, кн. ярослав. После него осталась дочь Мария, вышедшая замуж да смол. кн. Федора Ростиславича Чёрного и родившая ему сына Михаила — основателя рода кн. Ушатых, а также др. княж. родов: Моложских, Сицких, Прозоровских и пр., рано потерявших уделы и перешедших на службу в Москву.
Фонвизины — русский дворянский род,
происходивший от немцев из Ордена меченосцев. Один
из них, барон Петер фон Висин, вместе с сыном
Денисом был взят в плен войском Ивана IV Васильевича Грозного во время Ливонской
воины 1558 -1583. Они остались в России, куда
прибыли и младшие сыновья барона — Борис и Юрий.
Черкасские — княжеский род, слоджившийся в России во второй половине XVI века преимущественно из выехавших на русскую службу кабардинских вельмож — потомков Идара-Мурзы. Фамилия Черкасских произошла от укоренившегося в средневековой литературе названия северо-кавказских адыгейских племен — черкесов (черкасов — русских документах). Наиболее известные из князей Черкасских: Михаил (Салтанкул) Темрюкович и Борис Канбулатович — племянник Темрюка.
|
 Б. Хвостов, по прозвищу
Отяй. Один из его сыновей, Иван Федорович, по
прозвищу Ёрш, был постельничим Ивана III
Васильевича. Ф. И. Зубатый-Отяев был (1513)
наместником в Красном; С. Т. Отяев командовал (1543)
большим полком в Лит. походе.
Б. Хвостов, по прозвищу
Отяй. Один из его сыновей, Иван Федорович, по
прозвищу Ёрш, был постельничим Ивана III
Васильевича. Ф. И. Зубатый-Отяев был (1513)
наместником в Красном; С. Т. Отяев командовал (1543)
большим полком в Лит. походе. кн. Дмитрию Ивановичу
«с Ольгердовичи» и участвовавшего к
Куликовской битве (1380), после которой получил чин
окольничего. В XVI и XVII вв. Погожевы служили
полковыми головами и городовыми воеводами.
кн. Дмитрию Ивановичу
«с Ольгердовичи» и участвовавшего к
Куликовской битве (1380), после которой получил чин
окольничего. В XVI и XVII вв. Погожевы служили
полковыми головами и городовыми воеводами. От него и
пошли Полевы, в начале XVI века перешедшие на
службу в Москву.
От него и
пошли Полевы, в начале XVI века перешедшие на
службу в Москву. Его потомки
по воле великого князя стали именоваться
Александровыми-Протопоповыми. Можайские дети
боярские, Юрий и Иван Ивановичи Протопоповы,
записаны в Тысячной книге лучших дворян и детей
боярских под 1550. Елизарий Юрьевич Протопопов был
(1577) воеводой в судовой рати и убит (1577) под
Венденом.
Его потомки
по воле великого князя стали именоваться
Александровыми-Протопоповыми. Можайские дети
боярские, Юрий и Иван Ивановичи Протопоповы,
записаны в Тысячной книге лучших дворян и детей
боярских под 1550. Елизарий Юрьевич Протопопов был
(1577) воеводой в судовой рати и убит (1577) под
Венденом. См.
Генеалогическую таблицу: Род
графов Толстых
См.
Генеалогическую таблицу: Род
графов Толстых Ухтомы. От него пошли кн. Ухтомские служившие в Москве головами и
воеводами.
Ухтомы. От него пошли кн. Ухтомские служившие в Москве головами и
воеводами. Потомки их несколько поколений исповедовали
лютеранскую религию, что не мешало им верно
служить новой родине, тем более что они за это
получали поместья и др. пожалования.
Потомки их несколько поколений исповедовали
лютеранскую религию, что не мешало им верно
служить новой родине, тем более что они за это
получали поместья и др. пожалования.границ | Таксономическое и метаболическое несоответствие у древнего рода Streptomyces
Введение
Разнообразие и экология микроорганизмов формируют наш мир — от управления большей частью глобального круговорота элементов до посредничества в сложных межвидовых взаимодействиях, которые уравновешивают здоровье и болезнь (Falkowski et al. , 2008; Lynch and Pedersen, 2016). Достижения в области молекулярных и вычислительных методов позволили нам раскрыть полный генетический состав организмов по всему древу жизни и пролить свет на их общие и различные таксономии, образ жизни и метаболизм (Louca et al., 2018). Недавние исследования показали, что экология может влиять на метаболический потенциал (Chevrette et al., 2019) и поток генов между микроорганизмами (Smillie et al., 2011). Таким образом, исследование взаимодействия между экологией и метаболизмом необходимо для понимания ландшафтов приспособленности, влияющих на микробную эволюцию.
, 2008; Lynch and Pedersen, 2016). Достижения в области молекулярных и вычислительных методов позволили нам раскрыть полный генетический состав организмов по всему древу жизни и пролить свет на их общие и различные таксономии, образ жизни и метаболизм (Louca et al., 2018). Недавние исследования показали, что экология может влиять на метаболический потенциал (Chevrette et al., 2019) и поток генов между микроорганизмами (Smillie et al., 2011). Таким образом, исследование взаимодействия между экологией и метаболизмом необходимо для понимания ландшафтов приспособленности, влияющих на микробную эволюцию.
В бактериальных сообществах, а не в видах, как для большинства растений и животных, функциональные единицы таксономии часто определяются операционными таксономическими единицами (OTU). Это мотивировано отсутствием морфологически отличительных признаков и неотъемлемыми проблемами, связанными с концепциями видов бактерий.Поскольку ранние исследования Вёзе и других были сосредоточены на типировании последовательностей ДНК и молекулярной филогенетике, ген 16S рибосомной РНК (рРНК) стал основой микробной экологии и эволюции. Подавляющее большинство исследований используют этот локус в качестве маркера для группировки бактерий в OTU, часто разделяя таксоны на группы, обладающие идентичностью последовательности гена 16S рРНК выше 97% (Stegen et al., 2013; Ghoul and Mitri, 2016; Kim et al., 2017). Однако латеральный перенос генов, переменная скорость эволюции и зависимость от произвольных отсечек сильно влияют на способность гена 16S описывать общие геномные и метаболические характеристики.Несмотря на высокое таксономическое разнообразие, описываемое OTU, микробные сообщества из сходной экологии могут демонстрировать конвергенцию геномных признаков и функциональных маркеров (Farjalla et al., 2016). Кроме того, для контекста, ген 18S рРНК, эукариотическая малая субъединица рРНК со структурной и функциональной гомологией 16S у бактерий, является высококонсервативным внутри групп и может проявлять изменчивость, которая не обязательно соответствует эволюционным отношениям (рис. 1А). Непоследовательное присвоение таксономических уровней сбивает с толку бактериальное древо жизни; генетическое разнообразие «вида» Escherichia coli находится на уровне многих родов.
Подавляющее большинство исследований используют этот локус в качестве маркера для группировки бактерий в OTU, часто разделяя таксоны на группы, обладающие идентичностью последовательности гена 16S рРНК выше 97% (Stegen et al., 2013; Ghoul and Mitri, 2016; Kim et al., 2017). Однако латеральный перенос генов, переменная скорость эволюции и зависимость от произвольных отсечек сильно влияют на способность гена 16S описывать общие геномные и метаболические характеристики.Несмотря на высокое таксономическое разнообразие, описываемое OTU, микробные сообщества из сходной экологии могут демонстрировать конвергенцию геномных признаков и функциональных маркеров (Farjalla et al., 2016). Кроме того, для контекста, ген 18S рРНК, эукариотическая малая субъединица рРНК со структурной и функциональной гомологией 16S у бактерий, является высококонсервативным внутри групп и может проявлять изменчивость, которая не обязательно соответствует эволюционным отношениям (рис. 1А). Непоследовательное присвоение таксономических уровней сбивает с толку бактериальное древо жизни; генетическое разнообразие «вида» Escherichia coli находится на уровне многих родов. Некоторые роды могут охватывать сотни миллионов лет расхождения. Это мотивировало переопределение бактериального древа жизни на основе геномного сходства в общих локусах (Parks et al., 2018). Тем не менее, вариабельные области оказывают большое влияние на метаболизм и, таким образом, приспособленность организмов и метаболических единиц часто являются грузом латерального переноса (McDonald and Currie, 2017).
Некоторые роды могут охватывать сотни миллионов лет расхождения. Это мотивировало переопределение бактериального древа жизни на основе геномного сходства в общих локусах (Parks et al., 2018). Тем не менее, вариабельные области оказывают большое влияние на метаболизм и, таким образом, приспособленность организмов и метаболических единиц часто являются грузом латерального переноса (McDonald and Currie, 2017).
Рисунок 1 . (A) Идентичность с ближайшим совпадением генов 18S рРНК человека для нескольких многоклеточных животных.Центр, медиана; поле, верхний и нижний квантили; усы, 1,5-кратный межквартильный размах; баллы, выбросы. (B) Временная шкала, показывающая происхождение Streptomyces и основной раскол между Streptomyces Clades I и II в связи с другими интересными событиями. млн лет назад, млн лет назад; планки погрешностей, нижняя и верхняя оценки достоверности. Геологические периоды отмечены чередующимися цветами.
Как группа экологически и промышленно важный род Streptomyces представляет собой бактериальный таксон, обычно встречающийся в почве и микробиомах, связанных с хозяином (Hopwood, 2007; Andam et al., 2016; Шевретт и др., 2019). Возникнув примерно в тот же период времени, что и наземные растения (рис. 1B; McDonald and Currie, 2017), Streptomyces обладают разнообразными способностями к биосинтезу, а их вторичные метаболиты являются основным источником антибиотиков, используемых в клинической практике. Кроме того, Streptomyces могут продуцировать многие другие важные с экологической и медицинской точки зрения биоактивные молекулы и часто выделяют 15% или более своих геномов для вторичного метаболизма (Chevrette and Currie, 2019).Геномы Streptomyces в среднем имеют длину 8 мегабаз (Мб) (однако некоторые штаммы более чем на 25% больше) и примерно на 72% состоят из GC (Hopwood, 2007). Хотя их часто изучают как почвенные организмы, они легко связываются с разнообразными эукариотическими хозяевами, включая множество различных видов насекомых (Chevrette et al. , 2019). В пределах линий Streptomyces диверсификация метаболизма может совпадать с дивергенцией (Choudoir et al., 2018), а вариация метаболической способности может происходить даже при одинаковом 16S (Antony-Babu et al., 2017). Однако относительно мало известно о филогенетических и геномных влияниях на метаболизм в филогении Streptomyces .
, 2019). В пределах линий Streptomyces диверсификация метаболизма может совпадать с дивергенцией (Choudoir et al., 2018), а вариация метаболической способности может происходить даже при одинаковом 16S (Antony-Babu et al., 2017). Однако относительно мало известно о филогенетических и геномных влияниях на метаболизм в филогении Streptomyces .
Здесь мы исследуем несоответствие между таксономическим и метаболическим разнообразием у Streptomyces , чтобы оценить закономерности на нескольких филогенетических уровнях. В 288 готовых или почти готовых геномах Streptomyces мы исследуем взаимосвязь между коровым геномом, вариабельным геномом и идентичностью гена 16S рРНК с метаболическими и экологически значимыми генетическими элементами.В подгруппе из 28 штаммов, распределенных по филогении Streptomyces , мы описываем сдвиги в геномном содержании по отношению к экологии и характеризуем их экзометаболические профили.
Материалы и методы
Поиск данных
Все общедоступные геномы Streptomyces были получены из NCBI 31 августа 2018 г. , и в рамках этого исследования использовались геномы с восемью контигами или менее ( n = 259). 29 дополнительных геномов (28 Streptomyces и 1 Pseudonocardia внешней группы) были секвенированы и собраны в настоящем документе (см. ниже).Доступ к геномам можно получить по адресу https://bitbucket.org/chevrm/streptomyces_metabolites/.
, и в рамках этого исследования использовались геномы с восемью контигами или менее ( n = 259). 29 дополнительных геномов (28 Streptomyces и 1 Pseudonocardia внешней группы) были секвенированы и собраны в настоящем документе (см. ниже).Доступ к геномам можно получить по адресу https://bitbucket.org/chevrm/streptomyces_metabolites/.
Условия роста
пробки мицелиального агара (диаметром 1 мм) удаляли из активно растущих чашек YMEA с целевыми штаммами и инокулировали в 5 мл минимальной среды Хопвуда, содержащей 5% (вес/объем) глюкозы (пять повторов каждого штамма). Культуры инкубировали в течение 2 недель при 30°С на ротационном шейкере при 300 об/мин. Культуры центрифугировали при 10000× g в течение 10 мин, супернатанты собирали и хранили при –20°С.
Секвенирование и сборка
Культуры выращивали в богатой среде с добавлением 0,5% глицина, и клетки собирали центрифугированием. Клетки промывали 10,3%-ным раствором сахарозы, ресуспендировали в растворе лизоцима (3 мг/мл -1 лизоцима, Sigma, в 0,3 М сахарозе, 25 мМ Трис, рН 8 и 25 мМ ЭДТА, рН 8), и инкубировали при 37°С в течение 30 минут. Протеиназу К (Thermo Fisher; 20 мг/мл -1 ) добавляли перед инкубацией в течение 15 мин при 42°С. Клетки лизировали, добавляя 2% SDS и встряхивая в течение 5 мин до завершения лизиса.Добавляли нейтральный фенол и хлороформ и осторожно встряхивали пробирки до однородного белого цвета. После центрифугирования верхний слой переносили в 3 М раствор ацетата натрия, рН 6, и изопропанол. Пробирки осторожно перемешивали до появления ДНК. ДНК осаждали, супернатант удаляли и осадок ресуспендировали в ТЕ с 0,2 мг мл -1 РНКазы А. Пробирки инкубировали 15 мин при 28°C перед добавлением 5 М NaCl и раствора ЦТАБ/NaCl. Пробирки инкубировали 10 мин при 55°С и охлаждали до 28°С. Добавляли CHCl3, пробирки осторожно встряхивали и вращали в течение 10 мин при 28°C.Верхний слой переносили в новую пробирку и снова экстрагировали фенолом и хлороформом с последующей экстракцией хлороформом и осаждением 3 М ацетатом натрия, рН 6, и изопропанолом. Осадок промывали 70% этанолом и ресуспендировали в воде.
Протеиназу К (Thermo Fisher; 20 мг/мл -1 ) добавляли перед инкубацией в течение 15 мин при 42°С. Клетки лизировали, добавляя 2% SDS и встряхивая в течение 5 мин до завершения лизиса.Добавляли нейтральный фенол и хлороформ и осторожно встряхивали пробирки до однородного белого цвета. После центрифугирования верхний слой переносили в 3 М раствор ацетата натрия, рН 6, и изопропанол. Пробирки осторожно перемешивали до появления ДНК. ДНК осаждали, супернатант удаляли и осадок ресуспендировали в ТЕ с 0,2 мг мл -1 РНКазы А. Пробирки инкубировали 15 мин при 28°C перед добавлением 5 М NaCl и раствора ЦТАБ/NaCl. Пробирки инкубировали 10 мин при 55°С и охлаждали до 28°С. Добавляли CHCl3, пробирки осторожно встряхивали и вращали в течение 10 мин при 28°C.Верхний слой переносили в новую пробирку и снова экстрагировали фенолом и хлороформом с последующей экстракцией хлороформом и осаждением 3 М ацетатом натрия, рН 6, и изопропанолом. Осадок промывали 70% этанолом и ресуспендировали в воде. ДНК определяли количественно, проверяли на чистоту и пропускали через гель для проверки высокой молекулярной массы. Библиотеки геномной ДНК для секвенирования парных концов Illumina MiSeq 2 × 300 п.н. были подготовлены Биотехнологическим центром Университета Висконсин-Мэдисон (TruSeq).Чтения были исправлены с помощью MUSKET v1.1 (Liu et al., 2013), парные концы были объединены с помощью FLASH v1.2.7 (Magoč and Salzberg, 2011) и собраны с помощью SPAdes v3.11.0 (Bankevich et al., 2012). .
ДНК определяли количественно, проверяли на чистоту и пропускали через гель для проверки высокой молекулярной массы. Библиотеки геномной ДНК для секвенирования парных концов Illumina MiSeq 2 × 300 п.н. были подготовлены Биотехнологическим центром Университета Висконсин-Мэдисон (TruSeq).Чтения были исправлены с помощью MUSKET v1.1 (Liu et al., 2013), парные концы были объединены с помощью FLASH v1.2.7 (Magoč and Salzberg, 2011) и собраны с помощью SPAdes v3.11.0 (Bankevich et al., 2012). .
Геномная характеристика
последовательности генов 18S рРНК, аннотированные как Homo sapiens , были собраны из базы данных SILVA (Quast et al., 2012) и сопоставлены со всеми метазоями в SILVA с помощью blastn (Altschul et al., 1990). Лучшие несамостоятельные попадания в человека представлены на рисунке 1А. Временная шкала на рисунке 1B была создана на основе следующих оценок: происхождение H.sapiens 0,305 млн лет назад (млн лет назад; 0,26–0,35 млн лет назад низкая-высокая оценка) (Schlebusch et al.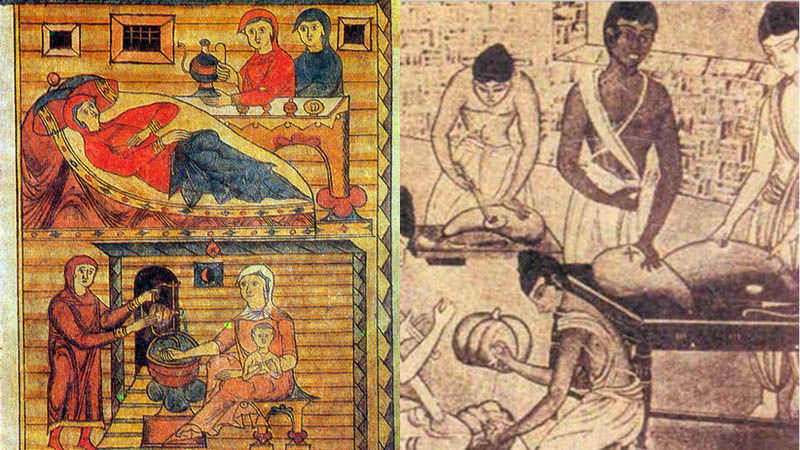 , 2017), происхождение приматов 74,1 млн лет назад (68,2–81,2 млн лет назад низкая-высокая оценка) (Pozzi et al., 2014) , происхождение наземных растений 457,5 млн лет назад (425–490 млн лет назад, низкая-высокая оценка) (Sanderson, 2003), происхождение Streptomyces 382 млн лет назад (250–514 млн лет назад, низкая-высокая оценка) (McDonald and Currie, 2017) и самая низкая общий предок основных Streptomyces Clades I и II 132 млн лет назад (низкая-высокая оценка 87–177 млн лет назад) (McDonald and Currie, 2017).
, 2017), происхождение приматов 74,1 млн лет назад (68,2–81,2 млн лет назад низкая-высокая оценка) (Pozzi et al., 2014) , происхождение наземных растений 457,5 млн лет назад (425–490 млн лет назад, низкая-высокая оценка) (Sanderson, 2003), происхождение Streptomyces 382 млн лет назад (250–514 млн лет назад, низкая-высокая оценка) (McDonald and Currie, 2017) и самая низкая общий предок основных Streptomyces Clades I и II 132 млн лет назад (низкая-высокая оценка 87–177 млн лет назад) (McDonald and Currie, 2017).
был создан с использованием 93 белков TIGRFAM в наборе основных бактериальных белков (GenProp0799; http://www.jcvi.org/cgi-bin/genome-properties/GenomePropDefinition.cgi?prop_acc=GenProp0799). Гены были вызваны с помощью prodigal v2.6.0 (Hyatt et al., 2010), и для поиска каждого генома использовались скрытые марковские модели профиля GenProp0799. HMMER v3.1b2 (Eddy, 2011) использовали для идентификации последовательностей белков для каждого семейства белков. Затем каждое семейство было сопоставлено с использованием MAFFT v7.245 (Katoh and Standley, 2013).Затем выравнивания были преобразованы в выравнивания кодонов и объединены. Многолокусная филогения была создана с использованием RAxML v8.1.24 (Stamatakis, 2014) в рамках модели замещения GTRgamma со 100 быстрыми бутстрапами. Дивергенция основного генома рассчитывалась как длина ветвей дерева генома, масштабированная от 0 (отсутствие расстояния) до 1 (наибольшее расстояние).
Затем каждое семейство было сопоставлено с использованием MAFFT v7.245 (Katoh and Standley, 2013).Затем выравнивания были преобразованы в выравнивания кодонов и объединены. Многолокусная филогения была создана с использованием RAxML v8.1.24 (Stamatakis, 2014) в рамках модели замещения GTRgamma со 100 быстрыми бутстрапами. Дивергенция основного генома рассчитывалась как длина ветвей дерева генома, масштабированная от 0 (отсутствие расстояния) до 1 (наибольшее расстояние).
гена 5S, 16S и 23S рРНК были предсказаны из геномов с barrnap v0.9 (Seemann, 2014, 2018). Предсказанные гены рРНК с длиной в пределах 20% от Streptomyces coelicolor 5S, 16S и 23S (117, 1738 и 3179 п.н. соответственно) были выровнены с mafft v7.310 (Katoh and Standley, 2013), и была рассчитана процентная идентичность нуклеотидов в столбцах выравнивания без пропусков между генами рРНК каждого генома. В геномах, где было предсказано более одного гена 5S, 16S или 23S рРНК, использовалось максимальное процентное совпадение идентичности.
гена были предсказаны с помощью prodigal v2.6.2 (Hyatt et al., 2010), а ортологи были предсказаны с помощью pyparanoid v0.3.1 (Melnyk et al., 2019). Функциональную характеристику ортологов проводили по лучшему битрейту DIAMOND blastp v0.9.24.125 (Buchfink et al., 2015) (покрытие запросов >60%) к базе данных Clusters of Orthologous Groups (COG) v2014 (Galperin et al., 2015). Элементы резистентности прогнозировались с помощью RGI из базы данных CARD v2.0.3 (McArthur et al., 2013). Семейства углеводных активных ферментов (CAZy) (Lombard et al., 2014) были идентифицированы с помощью HMMER v3.2.1 (Eddy, 2011) (пороговое значение e-значения 1e-5 для белков> 80aa, 1e-3 в противном случае). Метаболические признаки были идентифицированы с помощью скрытых марковских моделей профиля (pHMM) от Louca et al.(2018) и Anantharaman et al. (2016) в соответствии с требованиями присутствия/отсутствия в исследовании Louca. ANI между геномами был рассчитан с помощью fastANI (Jain et al., 2018). BGC были идентифицированы с помощью antiSMASH v4.0.2 (Blin et al., 2017) и сгруппированы в семейства кластеров генов (GCF) с помощью BiG-SCAPE (Navarro-Muñoz et al., 2018) (расстояние <0,3).
BGC были идентифицированы с помощью antiSMASH v4.0.2 (Blin et al., 2017) и сгруппированы в семейства кластеров генов (GCF) с помощью BiG-SCAPE (Navarro-Muñoz et al., 2018) (расстояние <0,3).
Для подмножества из 29 штаммов содержание общего гена было идентифицировано и визуализировано в anvi’o (Eren et al., 2015) через пайплайн пангенома (Delmont and Eren, 2018) (minbit = 0.5; mcl-инфляция = 4; минимальное появление = 2). Также для этих штаммов гены были кластеризованы с использованием OrthoMCL, реализованного в get_homologues (Contreras-Moreira and Vinuesa, 2013). Были введены пороговые значения кластеризации >75% охвата парного выравнивания и >75% процента идентичности аминокислотной последовательности. Дендрограмма UPGMA (метод невзвешенных парных групп со средним арифметическим) была сгенерирована из матрицы присутствия/отсутствия состава генома с подобием Жаккара и 100 повторениями начальной загрузки.
Частичное секвенирование ампликона 16S
Геномную ДНК выделяли из изолятов с использованием набора для выделения ДНК Powersoil в соответствии со спецификациями производителя (MoBio). Полимеразную цепную реакцию (ПЦР) проводили с использованием набора универсальных бактериальных праймеров 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′) и 1496 R (5′-CGG TTA CCT TGT TAC GAC TT-3′) из гипервариабельные области V1-V9. ПЦР проводили в стандартной реакции объемом 25 мкл с 1 мкл ДНК-матрицы (20 нг мкл -1 ), 12,5 мкл EconoTaq (корпорация Lucigen), 1 мкл комбинированных 10 мкМ праймеров и 12,5 мкл воды. За начальной денатурацией в течение 3 мин при 95°C следовал 3-минутный отжиг при 58°C, затем 35 циклов по 10 с при 96°C и 2 мин при 72°C, а также послецикловое удлинение при 72°C в течение 7 мин.Ампликоны подтверждали на 1,2% агарозном геле для электрофореза перед очисткой с помощью геля Wizard SV и системы ПЦР (Promega). Затем была проведена реакция секвенирования Big Dye с последующей вторичной очисткой перед отправкой на анализ в UW Biotech (Университет Висконсин-Мэдисон). Фрагменты гена 16S рРНК выравнивали с помощью SINA (Pruesse et al., 2012). Филогения была создана с использованием метода максимального правдоподобия и 500 повторов начальной загрузки в MEGA v6 (Tamura et al.
Полимеразную цепную реакцию (ПЦР) проводили с использованием набора универсальных бактериальных праймеров 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′) и 1496 R (5′-CGG TTA CCT TGT TAC GAC TT-3′) из гипервариабельные области V1-V9. ПЦР проводили в стандартной реакции объемом 25 мкл с 1 мкл ДНК-матрицы (20 нг мкл -1 ), 12,5 мкл EconoTaq (корпорация Lucigen), 1 мкл комбинированных 10 мкМ праймеров и 12,5 мкл воды. За начальной денатурацией в течение 3 мин при 95°C следовал 3-минутный отжиг при 58°C, затем 35 циклов по 10 с при 96°C и 2 мин при 72°C, а также послецикловое удлинение при 72°C в течение 7 мин.Ампликоны подтверждали на 1,2% агарозном геле для электрофореза перед очисткой с помощью геля Wizard SV и системы ПЦР (Promega). Затем была проведена реакция секвенирования Big Dye с последующей вторичной очисткой перед отправкой на анализ в UW Biotech (Университет Висконсин-Мэдисон). Фрагменты гена 16S рРНК выравнивали с помощью SINA (Pruesse et al., 2012). Филогения была создана с использованием метода максимального правдоподобия и 500 повторов начальной загрузки в MEGA v6 (Tamura et al. , 2013).
, 2013).
Экстракция и ЖХ-МС
Образцы замороженных сред (1 мл) лиофилизировали всухую, затем экстрагировали 200 мкл МеОН, содержащего 10 мкМ внутреннего стандарта (5–50 мкМ 13C, 15 N бесклеточная смесь аминокислот, #767964, Sigma), с последующим встряхиванием и водяной баней. обработка ультразвуком в течение 10 мин, снова встряхивание, затем обработка ультразвуком еще 5 мин.После центрифугирования при 5000 rcf в течение 5 минут супернатант удаляли и центрифугировали через мембрану 0,22 PVDF (Millipore, MultiScreen GV Filter Plate, кат. № MSGVS2210) в течение 3 минут при 2500 rcf в 96-луночный планшет. Экстракты высушивали в SpeedVac и хранили при -20°С.
При подготовке к масс-спектрометрии высушенные экстракты ресуспендировали в 150 мкл МеОН и обрабатывали ультразвуком на водяной бане. Тандемная масс-спектрометрия с жидкостной хроматографией (ЖХ-МС/МС) выполнялась для экстрактов с использованием комплекта ВЭЖХ Agilent 1290, при этом данные МС и МС/МС собирались с использованием Q Exactive Orbitrap MS (Thermo Scientific, Сан-Хосе, Калифорния). Полные спектры МС были собраны в диапазоне m/z 70–1050 при разрешении 70 000 FWHM, а данные фрагментации МС/МС были получены с использованием энергий столкновения 10, 20 и 30 В при разрешении 17 500 FWHM. Параметры прибора МС включали расход защитного газа 50 (а.е.), расход вспомогательного газа 20 (а.е.), расход продувочного газа 2 (а.е.), напряжение распыления 3 кВ и температуру капилляра 400°C. Нормально-фазовую хроматографию выполняли с использованием колонки HILIC (Millipore SeQuant ZIC-HILIC, 150 мм × 2,1 мм, 5 мкм, № по каталогу 50454) при 40°C и с использованием 2 мкл инжектируемого объема для каждого образца.Колонку уравновешивали 100% буфером B (95:5 ACN:H 2 O w/5 мМ ацетата аммония) в течение 1,5 мин при 0,45 мл/мин, разбавляя буфер B до 65% буфером A (H 2 O w/5 мМ ацетата аммония) в течение 13,5 мин, снижение до 0% В в течение 3 мин при увеличении потока до 0,6 мл/мин с последующей изократической элюцией в 100% буфере А в течение 5 мин. Метаболиты идентифицировали на основании точной массы и времени удерживания в сочетании со сравнением спектров фрагментации МС/МС с приобретенными стандартами.
Полные спектры МС были собраны в диапазоне m/z 70–1050 при разрешении 70 000 FWHM, а данные фрагментации МС/МС были получены с использованием энергий столкновения 10, 20 и 30 В при разрешении 17 500 FWHM. Параметры прибора МС включали расход защитного газа 50 (а.е.), расход вспомогательного газа 20 (а.е.), расход продувочного газа 2 (а.е.), напряжение распыления 3 кВ и температуру капилляра 400°C. Нормально-фазовую хроматографию выполняли с использованием колонки HILIC (Millipore SeQuant ZIC-HILIC, 150 мм × 2,1 мм, 5 мкм, № по каталогу 50454) при 40°C и с использованием 2 мкл инжектируемого объема для каждого образца.Колонку уравновешивали 100% буфером B (95:5 ACN:H 2 O w/5 мМ ацетата аммония) в течение 1,5 мин при 0,45 мл/мин, разбавляя буфер B до 65% буфером A (H 2 O w/5 мМ ацетата аммония) в течение 13,5 мин, снижение до 0% В в течение 3 мин при увеличении потока до 0,6 мл/мин с последующей изократической элюцией в 100% буфере А в течение 5 мин. Метаболиты идентифицировали на основании точной массы и времени удерживания в сочетании со сравнением спектров фрагментации МС/МС с приобретенными стандартами.
Данные ЖХ-МС были проанализированы с помощью ранее описанных методов (Yao et al., 2015). Набор критериев использовали для оценки каждого из обнаруженных метаболитов и присвоения уровня уверенности в идентификации на основе (1) абсолютного значения отличия времени удерживания от стандарта, (2) ошибки массы (ppm) обнаруженных m /z по сравнению с теоретическим m/z, и (3) показатель МС/МС, сравнивающий экспериментальный образец фрагментации МС/МС со стандартным. Для сравнения времени удерживания и МС/МС стандарты анализировали с использованием тех же методов ЖХ-МС, что и для образцов. Соединения с самым высоким уровнем положительной идентификации обнаруживали m/z <5 ppm от теоретического стандарта, <0.5-минутная разница во времени удерживания по сравнению со стандартом, а также сопоставление спектров фрагментации МС/МС либо с внешней базой данных (METLIN), либо с внутренней базой данных, созданной из стандартов, запущенных и собранных на МС Q Exactive Orbitrap.
Для каждого метаболита, присутствующего (относительно контрольных сред) как минимум в трех из пяти повторностей, содержание суммировалось как для положительных, так и для отрицательных ионов, и для последующего анализа использовалась самая высокая сумма. Затем геномы и коды INCHI для каждого штамма были обработаны с помощью MAGI (Erbilgin et al., 2019). Затем для каждого метаболита показатели MAGI для каждого метаболита нормализовали по шкале от 0 до 1 путем деления на показатель MAGI высшего штамма для данного метаболита. Дисперсионный анализ (ANOVA) пиковой интенсивности идентифицированных метаболитов проводили с использованием MetaboAnalyst 4.0 (Chong et al., 2018). Метаболиты с >50% отсутствующих значений удаляли. Оставшиеся пропущенные значения были заменены половиной минимального значения в исходных данных. Значения пиковой интенсивности были преобразованы с использованием обобщенной функции логарифмического преобразования и масштабированы с использованием функции автоматического масштабирования.Тепловая карта была создана с использованием евклидова расстояния и алгоритма кластеризации Уорда.
Затем геномы и коды INCHI для каждого штамма были обработаны с помощью MAGI (Erbilgin et al., 2019). Затем для каждого метаболита показатели MAGI для каждого метаболита нормализовали по шкале от 0 до 1 путем деления на показатель MAGI высшего штамма для данного метаболита. Дисперсионный анализ (ANOVA) пиковой интенсивности идентифицированных метаболитов проводили с использованием MetaboAnalyst 4.0 (Chong et al., 2018). Метаболиты с >50% отсутствующих значений удаляли. Оставшиеся пропущенные значения были заменены половиной минимального значения в исходных данных. Значения пиковой интенсивности были преобразованы с использованием обобщенной функции логарифмического преобразования и масштабированы с использованием функции автоматического масштабирования.Тепловая карта была создана с использованием евклидова расстояния и алгоритма кластеризации Уорда.
K Бломберга (Blomberg et al., 2003) по отношению к 16S, коровому геному и деревьям состава генов. Значимость ( p ) для каждой пары метаболит-дерево была протестирована с 1000 рандомизациями.
Значимость ( p ) для каждой пары метаболит-дерево была протестирована с 1000 рандомизациями.
Результаты
Взаимосвязь между идентичностью гена рРНК и филогенетической дивергенцией основного генома
Процентная идентичность нуклеотидов генов рРНК 16S, 23S и 5S была рассчитана между 288 штаммами (287 Streptomyces и 1 Pseudonocardia ) и сравнена с филогенетической дивергенцией основного генома (масштабированная длина ветви филогении из 93 консервативных бактериальных генов) (рис. 2А).Сходство гена 16S рРНК варьировалось от идентичного (100%) до примерно 88% ( Streptomyces против Pseudonocardia ). От 0 до ~ 0,15 дивергенции основного генома идентичность гена 16S рРНК падает со 100% до ~ 97%. Этот наклон уменьшается при расхождении от ~ 0,15 до ~ 0,30, где идентичность 16S остается относительно постоянной на уровне ~ 97–96%. Сравнение Streptomyces и Pseudonocardia (расхождение около 1,0) составляет ~ 88% идентичности гена 16S рРНК. Более длинный ген 23S рРНК имеет более крутой отрицательный наклон с меньшей идентичностью гена рРНК, чем более короткие 16S или 5S.5S, самый короткий показанный ген рРНК, имеет более высокую идентичность при низкой дивергенции основного генома, но более низкую идентичность при более высокой дивергенции. Несмотря на относительно единообразное представление дивергенции основного генома от 0 до ~0,6, все три гена рРНК имеют большое процентное падение на уровне ~0,4–0,5, 16S с ~93 до ~90%, 5S с ~94 до 85% и 23S с ~94 до 85%. от ~93 до ~90%, прежде чем снова упасть до ~85%.
Более длинный ген 23S рРНК имеет более крутой отрицательный наклон с меньшей идентичностью гена рРНК, чем более короткие 16S или 5S.5S, самый короткий показанный ген рРНК, имеет более высокую идентичность при низкой дивергенции основного генома, но более низкую идентичность при более высокой дивергенции. Несмотря на относительно единообразное представление дивергенции основного генома от 0 до ~0,6, все три гена рРНК имеют большое процентное падение на уровне ~0,4–0,5, 16S с ~93 до ~90%, 5S с ~94 до 85% и 23S с ~94 до 85%. от ~93 до ~90%, прежде чем снова упасть до ~85%.
Рисунок 2 . (A) Связь между дивергенцией основного генома и генами 5S, 16S и 23S рРНК у Streptomyces .Пунктирная линия = 97%; размер точки = количество попарных сравнений. (B) Связь между дивергенцией основного генома и общими семействами генов у Streptomyces . Размер точки = количество попарных сравнений. (C) Распределение семейств генов в филогении Streptomyces .
Распределение ортологов относительно филогенетической дивергенции основного генома
Внутри геномов было идентифицировано 64 000 генных семейств. Перекрытие в семействах генов (ортологи) относительно дивергенции основного генома имеет больший отрицательный наклон, чем гены рРНК (рис. 2В).Очень похожие основные геномы (т. Е. ~0 расхождений основного генома) в среднем имеют более 6000 общих ортологов, но это число быстро уменьшается до расхождения основного генома ~ 0,15. Здесь кривая принимает почти ступенчатую форму с плато на ~ 4500, ~ 4000 и ~ 3500 семейств генов, общих при дивергенции основного генома ~ 0, 15, ~ 0, 2 и ~ 0, 3 соответственно. Большинство семейств генов (51 888; 81% ортологов) обнаруживаются менее чем у 10% штаммов, и только 2520 семейств генов являются общими у 90% или более из 287 Streptomyces (рис. 2C).
Идентичность гена 16S рРНК в сравнении со средней идентичностью нуклеотидов и составом ортологов
Идентичность 16S не позволяет предсказать среднюю нуклеотидную идентичность (ANI) попарно выровненных областей (рис. 3A). Среднее значение ANI составило 81,69% и колебалось от 76,02 до 100%. При 100% идентичности нуклеотидов 16S штаммы Streptomyces могут иметь всего 84,42% ANI, а для Streptomyces ANI и идентичности 16S не наблюдается линейной зависимости (отношение 1:1 показано пунктирной диагональю, рисунок 3A).От идентичных последовательностей 16S до общего порога OTU 97% идентичности 16S (пунктирная линия) ANI может варьироваться от 100% до 78,30%, что лишь немного уже, чем 100–87,61% идентичности 16S. Streptomyces coelicolor A3(2) ( Sc ) из клады II филогенеза Streptomyces имел 82,02, 81,70 и 76,38% ANI по сравнению с S. albus J1074 ( Sa 9074 ), S. 07 ; Sa . griseus NBRC 13350 ( Sg ; Clade I) и Pseudonocardia sp.SID8383 (внешняя группа) соответственно (рис. 3А). Выровненные области (то есть общий генетический материал) в среднем составляли 4,12 Мб и могут варьироваться от 11,97 Мб до 0,93 Мб (рис. 3В). Геном Sg более выровнен с Sc (4,40 Мб), чем Sa — Sc (4,13 Мб), несмотря на то, что Sg и Sc принадлежат к разным линиям (клады I и II, соответственно; рис.
3A). Среднее значение ANI составило 81,69% и колебалось от 76,02 до 100%. При 100% идентичности нуклеотидов 16S штаммы Streptomyces могут иметь всего 84,42% ANI, а для Streptomyces ANI и идентичности 16S не наблюдается линейной зависимости (отношение 1:1 показано пунктирной диагональю, рисунок 3A).От идентичных последовательностей 16S до общего порога OTU 97% идентичности 16S (пунктирная линия) ANI может варьироваться от 100% до 78,30%, что лишь немного уже, чем 100–87,61% идентичности 16S. Streptomyces coelicolor A3(2) ( Sc ) из клады II филогенеза Streptomyces имел 82,02, 81,70 и 76,38% ANI по сравнению с S. albus J1074 ( Sa 9074 ), S. 07 ; Sa . griseus NBRC 13350 ( Sg ; Clade I) и Pseudonocardia sp.SID8383 (внешняя группа) соответственно (рис. 3А). Выровненные области (то есть общий генетический материал) в среднем составляли 4,12 Мб и могут варьироваться от 11,97 Мб до 0,93 Мб (рис. 3В). Геном Sg более выровнен с Sc (4,40 Мб), чем Sa — Sc (4,13 Мб), несмотря на то, что Sg и Sc принадлежат к разным линиям (клады I и II, соответственно; рис. 3B, соответственно; ). Sg — Sc имел больше невыровненного генома (8,45 Мб), чем Sa — Sc (7.27 Мб) (рис. 3С). При 100% идентичности 16S Streptomyces может иметь до 8,26 Мб невыровненного, вариабельного содержимого между штаммами, а при отсечении 97% 16S OTU это может увеличиться до 13,81 Мб (рис. 3C). Семейства общих генов варьировались от 921 до 2264, в среднем 2175,5 (рис. 3D). Большинство Streptomyces с более чем 94% идентичностью 16S имеют более 2000 семейств генов (рис. 3D), что согласуется с «общим» геномом Streptomyces (т. е. семействами генов, встречающимися более чем в 90% штаммов) 2520 ортологов (рис. 2C). ).В то время как идентичность 16S примерно коррелирует с общими ортологами (рис. 3D), почти идентичные штаммы по 16S (> 99% идентичности) могут иметь до 518 семейств неподеленных генов (рис. 3E). Корреляция 16S с наличием / отсутствием семейств генов, принадлежащих к функциональным категориям, включая вторичный метаболизм, демонстрирует аналогичные тенденции (дополнительные рисунки S1, S2).
3B, соответственно; ). Sg — Sc имел больше невыровненного генома (8,45 Мб), чем Sa — Sc (7.27 Мб) (рис. 3С). При 100% идентичности 16S Streptomyces может иметь до 8,26 Мб невыровненного, вариабельного содержимого между штаммами, а при отсечении 97% 16S OTU это может увеличиться до 13,81 Мб (рис. 3C). Семейства общих генов варьировались от 921 до 2264, в среднем 2175,5 (рис. 3D). Большинство Streptomyces с более чем 94% идентичностью 16S имеют более 2000 семейств генов (рис. 3D), что согласуется с «общим» геномом Streptomyces (т. е. семействами генов, встречающимися более чем в 90% штаммов) 2520 ортологов (рис. 2C). ).В то время как идентичность 16S примерно коррелирует с общими ортологами (рис. 3D), почти идентичные штаммы по 16S (> 99% идентичности) могут иметь до 518 семейств неподеленных генов (рис. 3E). Корреляция 16S с наличием / отсутствием семейств генов, принадлежащих к функциональным категориям, включая вторичный метаболизм, демонстрирует аналогичные тенденции (дополнительные рисунки S1, S2). Sg и Sc (Клада I против Клады II) имеют 2259 общих и только 5 неразделенных семейств генов, в то время как Sa — Sc (обе Клады II) имеют 2239 общих и 25 неразделенных семейств генов. Sc — P. sp. SID8383 имел 1170 общих и 1094 неразделенных семейства генов.
Sg и Sc (Клада I против Клады II) имеют 2259 общих и только 5 неразделенных семейств генов, в то время как Sa — Sc (обе Клады II) имеют 2239 общих и 25 неразделенных семейств генов. Sc — P. sp. SID8383 имел 1170 общих и 1094 неразделенных семейства генов.
Рисунок 3 . (A) Средняя идентичность нуклеотидов (ANI) по сравнению с идентичностью гена 16S рРНК при попарном сравнении Streptomyces . Диагональная линия = отношение 1:1. Длина (B) длины выравнивания ANI и (C) невыровненных частей обоих геномов по сравнению с идентичностью 16S в тех же парных сравнениях. Количество (D) общих и (E) неразделенных ортологов по сравнению с идентичностью 16S. (A–E) Вертикальная линия = 97% идентичности 16S; желтый = попарные сравнения также в подмножестве на рисунках 5, 6. Зеленый = клада II S. coelicolor A3(2) против клады II S. albus J1074; синий = ветвь II S. coelicolor A3(2) по сравнению с кладой I S. griseus NBRC 13350; красный = Clade II S. coelicolor A3(2) против Pseudonocardia sp. SID8383.
albus J1074; синий = ветвь II S. coelicolor A3(2) по сравнению с кладой I S. griseus NBRC 13350; красный = Clade II S. coelicolor A3(2) против Pseudonocardia sp. SID8383.
Метаболические и функциональные геномные особенности
Количество общих метаболических и функциональных геномных признаков, присутствующих у двух штаммов, сильно зависит от пороговых значений идентичности 16S (например,г., для OTU). Например, примерно два элемента, связанных с устойчивостью к антибиотикам, являются общими для штаммов со 100% идентичностью 16S (рис. 4А). Однако для штаммов Streptomyces , которые будут группироваться в пределах одной и той же OTU (т. е. > 97% идентичности 16S), среднее перекрытие элементов устойчивости падает почти до 0 (рис. 4А). Уникальные элементы сопротивления составляют ~ 2 или ~ 4 при 97% идентичности 16S для вызовов строгой и слабой достоверности соответственно (рис. 4А). Точно так же, когда кластеры биосинтетических генов (BGC) сгруппированы в семейства кластеров генов (GCF), происходит падение с ~ 21 до ~ 2 общих GCF со 100 до> 97% идентичности 16S (рис. 4B).Даже при 100% идентичности 16S в среднем 10 GCF являются уникальными и увеличиваются до более чем 50 при> 97% 16S (рис. 4B). Большинство GCF имеют мало членов, что указывает на то, что они редко встречаются в пределах Streptomyces , но некоторые из них встречаются в более чем 40 штаммах (рис. 4C). Большинство GCF ранее не были описаны, и в базе данных MIBiG нет совпадений BiG-SCAPE (Navarro-Muñoz et al., 2018) (рис. 4C; Medema et al., 2015). ~92 и ~79 общих семейств углеводно-активных ферментов (CAZy) (Lombard et al., 2014) наблюдаются при 100 и >97% 16S соответственно, в то время как ~5 и ~40 уникальных семейств CAZy наблюдаются при этих идентичностях 16S ( Рисунок 4D).Метаболические признаки (например, дыхание нитритов, восстановление арсената, уреаза и т. д.) варьируются от ~ 3,3 до ~ 3,0 общих признаков от 100 до> 97% 16S (рис. 4E). При 100% 16S в среднем нет различий в метаболических признаках между штаммами, в то время как при> 97% в среднем 0,6 являются уникальными (рис.
4B).Даже при 100% идентичности 16S в среднем 10 GCF являются уникальными и увеличиваются до более чем 50 при> 97% 16S (рис. 4B). Большинство GCF имеют мало членов, что указывает на то, что они редко встречаются в пределах Streptomyces , но некоторые из них встречаются в более чем 40 штаммах (рис. 4C). Большинство GCF ранее не были описаны, и в базе данных MIBiG нет совпадений BiG-SCAPE (Navarro-Muñoz et al., 2018) (рис. 4C; Medema et al., 2015). ~92 и ~79 общих семейств углеводно-активных ферментов (CAZy) (Lombard et al., 2014) наблюдаются при 100 и >97% 16S соответственно, в то время как ~5 и ~40 уникальных семейств CAZy наблюдаются при этих идентичностях 16S ( Рисунок 4D).Метаболические признаки (например, дыхание нитритов, восстановление арсената, уреаза и т. д.) варьируются от ~ 3,3 до ~ 3,0 общих признаков от 100 до> 97% 16S (рис. 4E). При 100% 16S в среднем нет различий в метаболических признаках между штаммами, в то время как при> 97% в среднем 0,6 являются уникальными (рис. 4E). Большинство штаммов Streptomyces демонстрируют метаболические признаки дыхания кислородом, уреазы и восстановления арсената, а более четверти штаммов демонстрируют признаки дыхания нитратов (рис. 4F).Окисление формальдегида, окисление метанола, фиксация углерода ( через Rubisco), нитратное дыхание и дыхание оксида азота у Streptomyces встречаются редко (рис. 4F).
4E). Большинство штаммов Streptomyces демонстрируют метаболические признаки дыхания кислородом, уреазы и восстановления арсената, а более четверти штаммов демонстрируют признаки дыхания нитратов (рис. 4F).Окисление формальдегида, окисление метанола, фиксация углерода ( через Rubisco), нитратное дыхание и дыхание оксида азота у Streptomyces встречаются редко (рис. 4F).
Рисунок 4 . Перекрытие между штаммами и уникальность между штаммами (A) резистентных элементов, (B) семейств кластеров биосинтетических генов (BGC), (D) семейств углеводно-активных ферментов и (E) метаболических признаков по сравнению с 16S пороги идентичности. (A) Цвета относятся к строгости совпадений с базой данных CARD, как неполных, так и строгих. (B) Цвета относятся либо к полным BGC (BGC с 0,5 kb или более фланкирующим контигом с обеих сторон), либо ко всем BGC, включая фрагменты. (C) Частота размеров семей для семейств генных кластеров. Цвета означают, имеет ли GCF соответствие MIBiG или ранее не было описано. (F) Фракция штаммов Streptomyces с каждым метаболическим признаком (с нулевым значением не показано).
(C) Частота размеров семей для семейств генных кластеров. Цвета означают, имеет ли GCF соответствие MIBiG или ранее не было описано. (F) Фракция штаммов Streptomyces с каждым метаболическим признаком (с нулевым значением не показано).
Геномная идентичность в источниках изолятов
Геномный состав подгруппы из 28 штаммов Streptomyces (показаны желтым цветом на рис. 3) и одного штамма Pseudonocardia (SID8383) был оценен и визуализирован in vi’o (рис. 5; Eren et al., 2015). Штаммы отмечены как связанные с насекомыми (синий), связанные с растениями (зеленый), связанные с грибами (фиолетовый) или почвенные (коричневый). Темное содержимое на каждой полосе карты генома означает наличие гена, а светлые области означают его отсутствие.Геномы упорядочены по охвату выравнивания ANI. Семь из 10 почвенных штаммов находятся в одной группе ANI. SID7817 (почва), SID4943 (связанный с насекомыми) и SID4952 (неизвестный источник) тесно связаны по охвату ANI, но демонстрируют различные генетические области, соответствующие рисунку 3. Основные группы почв, сгруппированные вместе, также демонстрируют переменное содержание. Между базовыми группами SID4943, SID1 и SID8360, связанными с насекомыми, можно увидеть лишь незначительные различия.
Основные группы почв, сгруппированные вместе, также демонстрируют переменное содержание. Между базовыми группами SID4943, SID1 и SID8360, связанными с насекомыми, можно увидеть лишь незначительные различия.
Рисунок 5 .Карта генома Anvi’o избранных Streptomyces . Каждый рельс принадлежит отдельному геному и окрашен в соответствии с его источником (синий = связан с насекомыми, зеленый = связан с растениями, фиолетовый = связан с грибами, почва = желто-коричневый, серый = неизвестно). Наличие (темное затенение) и отсутствие (светлое затенение) показано для всего содержания генов в двух или более штаммах. Штаммы упорядочены по охвату выравнивания ANI, а гены упорядочены по расстоянию присутствия/отсутствия (в обоих случаях евклидова кластеризация). Псевдонокардия SID8383 (внутренняя рейка) используется как внешняя группа.
Филогенетическое несоответствие метаболической способности
Частичный ген 16S рРНК (вариабельные области V1-V9), основной геном и состав генома (сходство Жаккара) приводят к противоречивым филогениям (рис. 6А). В то время как основной геном и состав наиболее похожи, в расположении кладов наблюдается мелкомасштабное несоответствие (см. зеленый, серый и SID4944, рисунок 6A). Частичный 16S дает филогению с низким разрешением (и часто неправильную), что подчеркивается большими политомиями и размещением SID4952 (серый, рисунок 6A).Наличие/отсутствие кластеризации GCF (рис. 6B) показывает некоторое, но не идеальное, соответствие основным группировкам генома. Аналогичные группировки наблюдаются в метаболических путях благодаря оценке ассоциации метаболитов и генов MAGI (Erbilgin et al., 2019), сочетающей геномику и метаболомику (рис. 6C, дополнительная фигура S3). В общей сложности 73 метаболита были произведены по-разному между штаммами (ANOVA, FDR <0,05; дополнительная фигура S3). Эти метаболиты сопоставлены с 63 путями KEGG у S. coelicolor (sco), 59 у S.avermitilis (sma) и 57 в S. griseus (sgr). Многие из этих метаболитов участвуют в метаболизме пиримидина, пурина, «аргинина и пролина», «глицина, серина и треонина».
6А). В то время как основной геном и состав наиболее похожи, в расположении кладов наблюдается мелкомасштабное несоответствие (см. зеленый, серый и SID4944, рисунок 6A). Частичный 16S дает филогению с низким разрешением (и часто неправильную), что подчеркивается большими политомиями и размещением SID4952 (серый, рисунок 6A).Наличие/отсутствие кластеризации GCF (рис. 6B) показывает некоторое, но не идеальное, соответствие основным группировкам генома. Аналогичные группировки наблюдаются в метаболических путях благодаря оценке ассоциации метаболитов и генов MAGI (Erbilgin et al., 2019), сочетающей геномику и метаболомику (рис. 6C, дополнительная фигура S3). В общей сложности 73 метаболита были произведены по-разному между штаммами (ANOVA, FDR <0,05; дополнительная фигура S3). Эти метаболиты сопоставлены с 63 путями KEGG у S. coelicolor (sco), 59 у S.avermitilis (sma) и 57 в S. griseus (sgr). Многие из этих метаболитов участвуют в метаболизме пиримидина, пурина, «аргинина и пролина», «глицина, серина и треонина».
Рисунок 6 . (A) Несогласующиеся отношения между Streptomyces по разным методам, частичное 16S (V3 и V4), филогения ядра генома или сходство состава генома по Жаккару (ортологи). (B) Наличие/отсутствие тепловой карты семейств кластеров биосинтетических генов (GCF) у Streptomyces .Строки = штаммы, столбцы = GCF. красный = присутствует; серый = отсутствует. (C) Показатели MAGI генома-метаболита (цвета; шкала от 0 до 1) в Streptomyces . Ряды = штаммы; столбцы = реакции; кластеризация = евклидова. (B,C) Полоса аннотаций для штаммов (цвета на рядовой дендрограмме) соответствуют кладам, обозначенным в (A) выше. (D) Филогенетический сигнал метаболитов по отношению к деревьям в (A) выше. Значимость отмечена p < 0.01 (слева от пунктирной линии).
Метаболиты были протестированы на соответствие топологиям 16S, основного генома и дерева состава генома на предмет филогенетического сигнала. Во всех случаях, когда метаболит достиг значимости ( p <0,01) по крайней мере для одной топологии дерева, по крайней мере одна другая топология не была значимой (рис. 6D). Например, филогенетический сигнал для аденина был значительным в дереве 16S, но не достиг значимости как в деревьях основного генома, так и в деревьях состава генома. Состав генома был относительно больше связан с метаболитами D-галактозамина, хинолин-4-карбоновой кислоты, ретиноевой кислоты и трегалозы.Уридин, лактоза и N,N-диметил-п-фенилендиамин лучше всего соответствуют основному геномному дереву. Другие метаболиты (например, L-пипеколиновая кислота, метилтиоаденозин, дипальмитоилцефалин) лучше всего соответствуют дереву 16S.
Во всех случаях, когда метаболит достиг значимости ( p <0,01) по крайней мере для одной топологии дерева, по крайней мере одна другая топология не была значимой (рис. 6D). Например, филогенетический сигнал для аденина был значительным в дереве 16S, но не достиг значимости как в деревьях основного генома, так и в деревьях состава генома. Состав генома был относительно больше связан с метаболитами D-галактозамина, хинолин-4-карбоновой кислоты, ретиноевой кислоты и трегалозы.Уридин, лактоза и N,N-диметил-п-фенилендиамин лучше всего соответствуют основному геномному дереву. Другие метаболиты (например, L-пипеколиновая кислота, метилтиоаденозин, дипальмитоилцефалин) лучше всего соответствуют дереву 16S.
Обсуждение
Близкородственные виды бактерий будут иметь генетическое и метаболическое перекрытие, обусловленное вертикальным наследованием генов (см. панели «перекрытия» на рисунках 4A, B, D, E). Однако давление отбора, которое формирует метаболические признаки и невертикальные механизмы генетического обмена, может привести к уникальным эволюционным историям. Некоторые черты могут быть сильно коррелированы с таксономией, а другие нет. Здесь мы количественно оцениваем, как эти общие характеристики коррелируют с общими мерами таксономической дистанции внутри Streptomyces . Чтобы сделать выводы об образе жизни, метаболизме и экологии микроорганизмов на основе геномных данных, требуется описание в более мелких филогенетических масштабах. Как область микробная экология в значительной степени построена на использовании гена 16S рРНК в качестве маркера для понимания бактериальной экологии и эволюции (Stegen et al., 2013; Гуль и Митри, 2016). Хотя ограничения 16S были признаны (Kim et al., 2017), мы считаем, что лучшее понимание и признание этих ограничений имеют решающее значение для этой области. Здесь мы оцениваем эти ограничения при описании взаимосвязи между метаболизмом, экологически важными наборами генов, коровым геномом и идентичностью 16S у Streptomyces . Посредством нашей оценки геномного и метаболического родства среди Streptomyces мы показываем, что признаки, которые способствуют метаболическому разнообразию, не полностью описываются общими группами таксономических единиц.
Некоторые черты могут быть сильно коррелированы с таксономией, а другие нет. Здесь мы количественно оцениваем, как эти общие характеристики коррелируют с общими мерами таксономической дистанции внутри Streptomyces . Чтобы сделать выводы об образе жизни, метаболизме и экологии микроорганизмов на основе геномных данных, требуется описание в более мелких филогенетических масштабах. Как область микробная экология в значительной степени построена на использовании гена 16S рРНК в качестве маркера для понимания бактериальной экологии и эволюции (Stegen et al., 2013; Гуль и Митри, 2016). Хотя ограничения 16S были признаны (Kim et al., 2017), мы считаем, что лучшее понимание и признание этих ограничений имеют решающее значение для этой области. Здесь мы оцениваем эти ограничения при описании взаимосвязи между метаболизмом, экологически важными наборами генов, коровым геномом и идентичностью 16S у Streptomyces . Посредством нашей оценки геномного и метаболического родства среди Streptomyces мы показываем, что признаки, которые способствуют метаболическому разнообразию, не полностью описываются общими группами таксономических единиц. В пределах Streptomyces идентичность 16S не коррелирует с наиболее основными экологически значимыми генами, такими как семейства CAZy и гены устойчивости к антибиотикам. Однако общий контент на концах дерева 16S (то есть почти идентичный по 16S) предполагает, что существуют группы Streptomyces с высококонсервативным содержанием генов, которые имеют отношение к экологии. В то время как 16S имеет некоторую полезность для прогнозирования грубых экологических признаков (например, почвенных и связанных с насекомыми в Chevrette et al., 2019) или таксономии на уровне рода, фенотипы и закономерности, которые проявляются в более мелких масштабах, могут оказаться неуловимыми даже при высокой идентичности 16S. .Это говорит о том, что если по 16S штаммы не идентичны, OTU не является значимой мерой экологически значимого разнообразия. Кроме того, высокая изменчивость даже в пределах идентичных последовательностей 16S предполагает, что будущие исследования должны отойти от основанных на 16S описаний экологии или метаболического потенциала.
В пределах Streptomyces идентичность 16S не коррелирует с наиболее основными экологически значимыми генами, такими как семейства CAZy и гены устойчивости к антибиотикам. Однако общий контент на концах дерева 16S (то есть почти идентичный по 16S) предполагает, что существуют группы Streptomyces с высококонсервативным содержанием генов, которые имеют отношение к экологии. В то время как 16S имеет некоторую полезность для прогнозирования грубых экологических признаков (например, почвенных и связанных с насекомыми в Chevrette et al., 2019) или таксономии на уровне рода, фенотипы и закономерности, которые проявляются в более мелких масштабах, могут оказаться неуловимыми даже при высокой идентичности 16S. .Это говорит о том, что если по 16S штаммы не идентичны, OTU не является значимой мерой экологически значимого разнообразия. Кроме того, высокая изменчивость даже в пределах идентичных последовательностей 16S предполагает, что будущие исследования должны отойти от основанных на 16S описаний экологии или метаболического потенциала.
Появление секвенирования 16S привело к беспрецедентно глубоким описаниям микробной среды (Moyer et al., 1994; Ferris et al., 1996; Hugenholtz et al., 1998; Gao et al., 2007; Grice et al., 2009) и даже привело к открытию архей (Woese and Fox, 1977). Эти исследования обеспечили филогенетическую основу, на основе которой мы определяем эволюционные отношения в микробной жизни. Однако изменчивость процессов на уровне видов и популяций (например, специализированный метаболизм, специализация ниш и т. д.) не разрешается идентичностью 16S, поскольку скорость дивергенции внутри 16S не обязательно коррелирует с геномным разнообразием под влиянием незафиксированных селективных факторов. давление. По аналогии, при отсечке 18S в 97% большинство людей, оленей и дельфинов будут сгруппированы в одну и ту же OTU (рис. 1А).Разнообразие внутри Streptomyces , представленное здесь, предполагает, что мы не можем сделать вывод об образе жизни, функции или экологическом фенотипе по таксономическим маркерам.
Заявление о доступности данных
Наборы данных, созданные для этого исследования, и версии каждой фигуры в высоком разрешении можно найти на странице https://bitbucket.org/chevrm/streptomyces_metabolites/src.
Вклад авторов
MC и CC-S выполнили геномный анализ и метаболомный анализ. MC, CC-S и CC написали рукопись.KL, BB и TN проводили метаболические эксперименты.
Финансирование
Этот проект был поддержан Национальными институтами здравоохранения (NIH) U19 Al109673 и NIH U19 TW009872. Дополнительная поддержка была оказана MC через награду Национальной исследовательской службы NIH T32 GM008505. Финансирование CC и MC было предоставлено Исследовательским центром биоэнергетики Великих озер, Министерством энергетики США, Управлением науки, Управлением биологических и экологических исследований под номерами DE-SC0018409 и DE-FC02-07ER64494.Работа, проводимая Объединенным институтом генома Министерства энергетики США, подразделением Министерства энергетики США по научным исследованиям, поддерживается Управлением науки Министерства энергетики США по контракту № DE-AC02-05Ch21231.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02170/full#supplementary-material
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S1 | Количество общих ортологов с различной идентичностью генов 16S рРНК в попарных сравнениях Streptomyces , стратифицированных по функциональной категории COG. Желтые — попарные сравнения также в подмножестве с рис. 5, 6; зеленый = клада II S. coelicolor A3(2) по сравнению с кладой II S. albus J1074; синий = Clade II S. coelicolor A3(2) vs.Клада I S. griseus NBRC 13350; красный = Clade II S. coelicolor A3(2) против Pseudonocardia sp. SID8383. [ (C) производство и преобразование энергии; (D) контроль клеточного цикла, клеточное деление и разделение хромосом; (E) транспорт и метаболизм аминокислот; (F) транспорт и метаболизм нуклеотидов; (G) транспорт и метаболизм углеводов; (H) транспорт и метаболизм коферментов; (I) транспорт и метаболизм липидов; (J) трансляция, рибосомная структура и биогенез; (К) транскрипция; (L) репликация, рекомбинация и репарация; (M) биогенез клеточной стенки/мембраны/оболочки; (N) подвижность клеток; (O) посттрансляционная модификация, белковый оборот и шапероны; (P) транспорт и метаболизм неорганических ионов; (Q) биосинтез, транспорт и катаболизм вторичных метаболитов; (R) только предсказание общей функции; (S) функция неизвестна; (T) механизмы передачи сигнала; (U) внутриклеточный транспорт, секреция и везикулярный транспорт; (V) защитные механизмы; (W) внеклеточных структур].
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S2 | Количество уникальных ортологов с различной идентичностью генов 16S рРНК в попарных сравнениях Streptomyces , стратифицированных по функциональной категории COG. Желтые — попарные сравнения также в подмножестве с рис. 5, 6; зеленый = клада II S. coelicolor A3(2) по сравнению с кладой II S. albus J1074; синий = ветвь II S. coelicolor A3(2) по сравнению с кладой I S. griseus NBRC 13350; красный = Клада II S.coelicolor A3(2) по сравнению с Pseudonocardia sp. SID8383. [ (C) производство и преобразование энергии; (D) контроль клеточного цикла, клеточное деление, разделение хромосом; (E) транспорт и метаболизм аминокислот; (F) транспорт и метаболизм нуклеотидов; (G) транспорт и метаболизм углеводов; (H) транспорт и метаболизм коферментов; (I) транспорт и метаболизм липидов; (J) трансляция, рибосомная структура и биогенез; (К) транскрипция; (L) репликация, рекомбинация и репарация; (M) биогенез клеточной стенки/мембраны/оболочки; (N) подвижность клеток; (O) посттрансляционная модификация, белковый оборот и шапероны; (P) Транспорт и метаболизм неорганических ионов; (Q) биосинтез, транспорт и катаболизм вторичных метаболитов; (R) только предсказание общей функции; (S) функция неизвестна; (T) механизмы передачи сигнала; (U) внутриклеточный транспорт, секреция и везикулярный транспорт; (V) защитные механизмы; (W) внеклеточных структур].
ДОПОЛНИТЕЛЬНЫЙ РИСУНОК S3 | Тепловая карта средней пиковой интенсивности для дифференциальных метаболитов на основе ANOVA (FDR <0,05). Тепловая карта была создана с использованием евклидова расстояния и алгоритма кластеризации Уорда.
Ссылки
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Базовый инструмент локального поиска выравнивания. Дж. Мол. биол. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Полнотекстовая перекрестная ссылка | Академия Google
Анантараман, К., Brown, C.T., Hug, L.A., Sharon, I., Castelle, C.J., Probst, A.J., et al. (2016). Тысячи микробных геномов проливают свет на взаимосвязанные биогеохимические процессы в системе водоносных горизонтов. Нац. коммун. 7, 1–11. doi: 10.1038/ncomms13219
Полнотекстовая перекрестная ссылка | Академия Google
Андам, С. П., Дорогази, Дж. Р., Кэмпбелл, А. Н., Келли, П. Дж., Шудуар, М. Дж., и Бакли, Д. Х. (2016). Градиент широтного разнообразия наземных бактерий рода Streptomyces . MBio 7, e02200–e02215. doi: 10.1128/mBio.02200-15
Х. (2016). Градиент широтного разнообразия наземных бактерий рода Streptomyces . MBio 7, e02200–e02215. doi: 10.1128/mBio.02200-15
Полнотекстовая перекрестная ссылка | Академия Google
Энтони-Бабу, С., Стьен, Д., Эпарвье, В., Паррот, Д., Томаси, С., и Судзуки, М. Т. (2017). Несколько видов Streptomyces с различными вторичными метаболомами имеют идентичные последовательности гена 16S рРНК. науч. Респ. 7, 1–8. doi: 10.1038/s41598-017-11363-1
Полнотекстовая перекрестная ссылка | Академия Google
Банкевич А., Нурк С., Антипов Д., Гуревич А.А., Дворкин М., Куликов А.С., и соавт. (2012). SPAdes: новый алгоритм сборки генома и его приложения для секвенирования отдельных клеток. Дж. Вычисл. биол. 19, 455–477. doi: 10.1089/cmb.2012.0021
Полнотекстовая перекрестная ссылка | Академия Google
Блин, К., Вольф, Т., Шевретт, М.Г., Лу, X., Швален, С.Дж., Каутсар, С.А., и соавт. (2017). antiSMASH 4.0 — улучшения в предсказании химии и идентификации границ кластера генов. Рез. нуклеиновых кислот. 45, W36–W41.doi: 10.1093/nar/gkx319
Рез. нуклеиновых кислот. 45, W36–W41.doi: 10.1093/nar/gkx319
Полнотекстовая перекрестная ссылка | Академия Google
Бломберг, С.П., Гарланд, Т., и Айвз, А.Р. (2003). Проверка на филогенетический сигнал в сравнительных данных: поведенческие признаки более лабильны. Эволюция 57, 717–745. doi: 10.1111/j.0014-3820.2003.tb00285.x
Полнотекстовая перекрестная ссылка | Академия Google
Бухфинк Б., Се К. и Хьюсон Д. Х. (2015). Быстрое и чувствительное выравнивание белков с помощью DIAMOND. Нац. Методы 12, 59–60.doi: 10.1038/nmeth.3176
Полнотекстовая перекрестная ссылка | Академия Google
Chevrette, M.G., Carlson, C.M., Ortega, H.E., Thomas, C., Ananiev, G.E., Barns, K.J., et al. (2019). Антимикробный потенциал Streptomyces из микробиомов насекомых. Нац. коммун. 10:516. doi: 10.1038/s41467-019-08438-0
Полнотекстовая перекрестная ссылка | Академия Google
Шевретт, М. Г., и Карри, Ч. Р. (2019). Новые эволюционные парадигмы в открытии антибиотиков. J. Ind. Microbiol. Биотехнолог. 46, 257–271. doi: 10.1007/s10295-018-2085-6
Р. (2019). Новые эволюционные парадигмы в открытии антибиотиков. J. Ind. Microbiol. Биотехнолог. 46, 257–271. doi: 10.1007/s10295-018-2085-6
Полнотекстовая перекрестная ссылка | Академия Google
Chong, J., Soufan, O., Li, C., Caraus, I., Li, S., Bourque, G., et al. (2018). MetaboAnalyst 4.0: к более прозрачному и комплексному анализу метаболомики. Рез. нуклеиновых кислот. 46, W486–W494. doi: 10.1093/nar/gky310
Полнотекстовая перекрестная ссылка | Академия Google
Шудуар, М., Пепе-Ранни, К., и Бакли, Д. (2018).Диверсификация кластеров генов биосинтеза вторичных метаболитов совпадает с дивергенцией клонов у Streptomyces . Антибиотики 7:12. doi: 10.3390/антибиотики7010012
Полнотекстовая перекрестная ссылка | Академия Google
Контрерас-Морейра, Б., и Винуэса, П. (2013). GET_HOMOLOGUES, универсальный программный пакет для масштабируемого и надежного анализа микробного пангенома. Заяв. Окружающая среда. микробиол. 79, 7696–7701. doi: 10.1128/AEM.02411-13
микробиол. 79, 7696–7701. doi: 10.1128/AEM.02411-13
Полнотекстовая перекрестная ссылка | Академия Google
Дельмонт, Т.О. и Эрен А.М. (2018). Связывание пангеномов и метагеномов: метапангеном Prochlorococcus . PeerJ 6:e4320. doi: 10.7717/peerj.4320
Полнотекстовая перекрестная ссылка | Академия Google
Erbilgin, O., Rübel, O., Louie, K.B., Trinh, M., de Raad, M., Wildish, T., et al. (2019). MAGI: метод метаболита, аннотации и интеграции генов. ACS Хим. биол. 14:204362. doi: 10.1021/acschembio.8b01107
Полнотекстовая перекрестная ссылка | Академия Google
Эрен, А.М., Эсен, О. C., Quince, C., Vineis, J.H., Morrison, H.G., Sogin, M.L., et al. (2015). Anvi’o: продвинутая платформа анализа и визуализации данных omics. PeerJ 3:e1319. doi: 10.7717/peerj.1319
Полнотекстовая перекрестная ссылка | Академия Google
Falkowski, P.G., Fenchel, T., and Delong, E.F. (2008). Микробные двигатели, управляющие биогеохимическими циклами Земли. Наука 320, 1034–1039. doi: 10.1126/science.1153213
Наука 320, 1034–1039. doi: 10.1126/science.1153213
Полнотекстовая перекрестная ссылка | Академия Google
Фарджалла, В.F., Doebeli, M., Jacques, S.M.S., Parfrey, L.W., Srivastava, D.S., Louca, S., et al. (2016). Высокая таксономическая изменчивость, несмотря на стабильную функциональную структуру микробных сообществ. Нац. Экол. Эвол. 1:15. doi: 10.1038/s41559-016-0015
Полнотекстовая перекрестная ссылка | Академия Google
Феррис М.Дж., Мюзер Г. и Уорд Д.М. (1996). Профили денатурирующего градиентного гель-электрофореза 16S рРНК-определяемых популяций, населяющих сообщество микробных матов горячих источников. Заяв. Окружающая среда. микробиол. 62, 340–346. Доступно по адресу: http://www.ncbi.nlm.nih.gov/pubmed/8593039
.Академия Google
Гальперин М.Ю., Макарова К.С., Вольф Ю.И., Кунин Е.В. (2015). Расширенный охват микробного генома и улучшенная аннотация семейства белков в базе данных COG. Рез. нуклеиновых кислот. 43, Д261–Д269. doi: 10.1093/nar/gku1223
43, Д261–Д269. doi: 10.1093/nar/gku1223
Полнотекстовая перекрестная ссылка | Академия Google
Гао З., Ценг К., Пей З. и Блазер М.Дж. (2007). Молекулярный анализ бактериальной биоты поверхностной кожи предплечья человека. Проц. Натл. акад. науч. США 104, 2927–2932. doi: 10.1073/pnas.0607077104
Полнотекстовая перекрестная ссылка | Академия Google
Гуль, М., и Митри, С. (2016). Экология и эволюция микробной конкуренции. Тенденции микробиол. 24, 833–845. doi: 10.1016/j.tim.2016.06.011
Полнотекстовая перекрестная ссылка | Академия Google
Грайс, Э. А., Конг, Х. Х., Конлан, С., Деминг, К.Б., Дэвис Дж., Янг А.С. и соавт. (2009). Топографическое и временное разнообразие микробиома кожи человека. Наука 324, 1190–1192. doi: 10.1126/science.1171700
Полнотекстовая перекрестная ссылка | Академия Google
Хопвуд, Д. А. (2007). Streptomyces в природе и медицине: создатели антибиотиков . Нью-Йорк: Издательство Оксфордского университета.
Академия Google
Hyatt, D., Chen, G.-L., Locascio, P.F., Land, M.L., Larimer, F.W., and Hauser, L.Дж. (2010). Prodigal: распознавание прокариотических генов и идентификация сайта инициации трансляции. Биоинформатика BMC 11:119. дои: 10.1186/1471-2105-11-119
Полнотекстовая перекрестная ссылка | Академия Google
Джайн, К., Родригес-Р., Л.М., Филлиппи, А.М., Константинидис, К.Т., и Алуру, С. (2018). Высокопроизводительный ANI-анализ 90 000 геномов прокариот выявляет четкие границы видов. Нац. коммун. 9:5114. doi: 10.1038/s41467-018-07641-9
Полнотекстовая перекрестная ссылка | Академия Google
Като, К.и Стэндли, Д.М. (2013). Программное обеспечение MAFFT для множественного выравнивания последовательностей, версия 7: улучшения производительности и удобства использования. Мол. биол. Эвол. 30, 772–780. doi: 10.1093/molbev/mst010
Полнотекстовая перекрестная ссылка | Академия Google
Kim, D. , Hofstaedter, C.E., Zhao, C., Mattei, L., Tanes, C., Clarke, E., et al. (2017). Оптимизация методов и избегание ловушек в исследованиях микробиома. Микробиом 5:52. doi: 10.1186/s40168-017-0267-5
, Hofstaedter, C.E., Zhao, C., Mattei, L., Tanes, C., Clarke, E., et al. (2017). Оптимизация методов и избегание ловушек в исследованиях микробиома. Микробиом 5:52. doi: 10.1186/s40168-017-0267-5
Полнотекстовая перекрестная ссылка | Академия Google
Лю, Ю., Шредер, Дж., и Шмидт, Б. (2013). Musket: многоступенчатый корректор ошибок на основе спектра k-mer для данных последовательности Illumina. Биоинформатика 29, 308–315. doi: 10.1093/биоинформатика/bts690
Полнотекстовая перекрестная ссылка | Академия Google
Ломбард В., Голаконда Рамулу Х., Друла Э., Коутиньо П. М. и Хенриссат Б. (2014). База данных углеводно-активных ферментов (CAZy) в 2013 г. Nucleic Acids Res. 42, Д490–Д495. doi: 10.1093/nar/gkt1178
Полнотекстовая перекрестная ссылка | Академия Google
Лука, С., Martiny, A.C., Polz, M.F., Doolittle, W.F., Nelson, M.B., Hahn, A.S., et al. (2018). Функция и функциональная избыточность в микробных сообществах. Нац. Экол. Эвол. 2, 936–943. doi: 10.1038/s41559-018-0519-1
Нац. Экол. Эвол. 2, 936–943. doi: 10.1038/s41559-018-0519-1
Полнотекстовая перекрестная ссылка | Академия Google
Линч С.В. и Педерсен О. (2016). Микробиом кишечника человека в норме и при патологии. Н. англ. Дж. Мед. 375, 2369–2379. doi: 10.1056/NEJMra1600266
Полнотекстовая перекрестная ссылка | Академия Google
Магоч, Т.и Зальцберг С.Л. (2011). FLASH: быстрая корректировка длины коротких ридов для улучшения сборки генома. Биоинформатика 27, 2957–2963. doi: 10.1093/биоинформатика/btr507
Полнотекстовая перекрестная ссылка | Академия Google
Макартур, А. Г., Ваглехнер, Н., Низам, Ф., Ян, А., Азад, М. А., Бейлай, А. Дж., и соавт. (2013). Полная база данных по устойчивости к антибиотикам. Антимикроб. Агенты Чемотер. 57, 3348–3357. doi: 10.1128/AAC.00419-13
Полнотекстовая перекрестная ссылка | Академия Google
Макдональдс, Б.Р. и Карри, Ч.Р. (2017). Динамика латерального переноса генов у древних бактерий рода Streptomyces . МБио 8: e00644–17. doi: 10.1128/mBio.00644-17
МБио 8: e00644–17. doi: 10.1128/mBio.00644-17
Полнотекстовая перекрестная ссылка | Академия Google
Медема, М. Х., Коттманн, Р., Йилмаз, П., Каммингс, М., Биггинс, Дж. Б., Блин, К., и соавт. (2015). Минимум информации о кластере биосинтетических генов. Нац. хим. биол. 11, 625–631. doi: 10.1038/nchembio.1890
Полнотекстовая перекрестная ссылка | Академия Google
Мельник, Р.А., Хоссейн С.С. и Хейни С.Х. (2019). Конвергентное приобретение и потеря геномных островов вызывают изменения образа жизни у связанных с растениями Pseudomonas . ISME J. 13, 704–714. doi: 10.1038/s41396-019-0372-5
Полнотекстовая перекрестная ссылка | Академия Google
Мойер, К.Л., Доббс, Ф.К., и Карл, Д.М. (1994). Оценка разнообразия и структуры сообщества с помощью анализа распределения полиморфизма длин рестрикционных фрагментов бактериальных генов 16S рРНК из микробного мата в активной системе гидротермальных источников, подводная гора Лоихи, Гавайи. Заяв. Окружающая среда. микробиол. 60, 871–879. Доступно по адресу: http://www.ncbi.nlm.nih.gov/pubmed/7512808
Заяв. Окружающая среда. микробиол. 60, 871–879. Доступно по адресу: http://www.ncbi.nlm.nih.gov/pubmed/7512808
Академия Google
Наварро-Муньос, Дж., Селем-Мохика, Н., Маллоуни, М., Каутсар, С., Трион, Дж., Паркинсон, Э., и соавт. (2018). Вычислительная структура для систематического исследования биосинтетического разнообразия на основе крупномасштабных геномных данных. bioRxiv [Препринт]:445270. дои: 10.1101/445270
Полнотекстовая перекрестная ссылка | Академия Google
Паркс, Д.Х., Чувочина М., Уэйт Д.В., Ринке С., Скаршевски А., Шомейл П. и соавт. (2018). Стандартизированная таксономия бактерий, основанная на филогении генома, существенно пересматривает древо жизни. Нац. Биотехнолог. 36, 996–1004. doi: 10.1038/nbt.4229
Полнотекстовая перекрестная ссылка | Академия Google
Поцци, Л., Баррелл, А.С., Стернер, К.Н., Ходжсон, Дж.А., Дисотелл, Т.Р., и Раум, Р.Л. (2014). Филогенетические отношения приматов и даты расхождения, полученные на основе полных митохондриальных геномов. Мол. Филогенет. Эвол. 75, 165–183. doi: 10.1016/j.ympev.2014.02.023
Мол. Филогенет. Эвол. 75, 165–183. doi: 10.1016/j.ympev.2014.02.023
Полнотекстовая перекрестная ссылка | Академия Google
Pruesse, E., Peplies, J., и Glöckner, F. O. (2012). SINA: точное высокопроизводительное множественное выравнивание последовательностей генов рибосомных РНК. Биоинформатика 28, 1823–1829. doi: 10.1093/биоинформатика/bts252
Полнотекстовая перекрестная ссылка | Академия Google
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al.(2012). Проект базы данных генов рибосомной РНК SILVA: улучшенная обработка данных и веб-инструменты. Рез. нуклеиновых кислот. 41, Д590–Д596. doi: 10.1093/nar/gks1219
Полнотекстовая перекрестная ссылка | Академия Google
Сандерсон, MJ (2003). Молекулярные данные по 27 белкам не подтверждают докембрийское происхождение наземных растений. утра. Дж. Бот. 90, 954–956. doi: 10.3732/ajb.90.6.954
Полнотекстовая перекрестная ссылка | Академия Google
Schlebusch, C. M., Malmström, H., Günther, T., Sjödin, P., Coutinho, A., Edlund, H., et al. (2017). Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад. Наука 358, 652–655. doi: 10.1126/science.aao6266
M., Malmström, H., Günther, T., Sjödin, P., Coutinho, A., Edlund, H., et al. (2017). Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад. Наука 358, 652–655. doi: 10.1126/science.aao6266
Полнотекстовая перекрестная ссылка | Академия Google
Зееманн, Т. (2014). Prokka: быстрая аннотация генома прокариот. Биоинформатика 30, 2068–2069. doi: 10.1093/биоинформатика/btu153
Полнотекстовая перекрестная ссылка | Академия Google
Смайли, К.С., Смит, М.Б., Фридман, Дж., Кордеро, О.К., Дэвид, Л.А., и Алм, Э.Дж. (2011). Экология управляет глобальной сетью обмена генами, соединяющей человеческий микробиом. Природа 480, 241–244. doi: 10.1038/nature10571
Полнотекстовая перекрестная ссылка | Академия Google
Стаматакис, А. (2014). RAxML версии 8: инструмент для филогенетического анализа и пост-анализа больших филогений. Биоинформатика 30, 1312–1313. doi: 10. 1093/биоинформатика/btu033
1093/биоинформатика/btu033
Полнотекстовая перекрестная ссылка | Академия Google
Стеген, Дж.C., Lin, X., Fredrickson, J.K., Chen, X., Kennedy, D.W., Murray, C.J., et al. (2013). Количественная оценка процессов сборки сообщества и определение особенностей, которые их налагают. ISME J. 7, 2069–2079. doi: 10.1038/ismej.2013.93
Полнотекстовая перекрестная ссылка | Академия Google
Тамура К., Стечер Г., Петерсон Д., Филипски А. и Кумар С. (2013). MEGA6: молекулярно-эволюционный генетический анализ версии 6.0. Мол. биол. Эвол. 30, 2725–2729. doi: 10.1093/molbev/mst197
Полнотекстовая перекрестная ссылка | Академия Google
Везе, К.Р. и Фокс, Г.Е. (1977). Филогенетическая структура прокариотического домена: первичные царства. Проц. Натл. акад. науч. США 74, 5088–5090. doi: 10.1073/pnas.74.11.5088
Полнотекстовая перекрестная ссылка | Академия Google
Яо Ю., Сунь Т., Ван Т., Рюбель О., Нортен Т. и Боуэн Б. (2015). Анализ наборов метаболомных данных с помощью высокопроизводительных вычислений и атласов метаболитов. Мета 5, 431–442. doi: 10.3390/metabo5030431
и Боуэн Б. (2015). Анализ наборов метаболомных данных с помощью высокопроизводительных вычислений и атласов метаболитов. Мета 5, 431–442. doi: 10.3390/metabo5030431
Полнотекстовая перекрестная ссылка | Академия Google
Розничная цена для студентов: 25 долларов.00 Джоанна Брюк 9780192858252 | |
Третье издание Розничная цена для студентов: 69,95 $ Ян Моррис и Барри Б. Пауэлл 9780197586891 | |
Второе издание Розничная цена для студентов: 18 долларов.95 Грег Вульф 97801 458Мягкая обложка Сентябрь 2021 г. | |
Розничная цена для студентов: 24,95 $ Селия Э. 9780197601839 | |
Розничная цена для студентов: 24 доллара.95 Натанаэль Андраде 9780197610817 | |
Розничная цена для студентов: 24,95 $ Хайди Маркс 9780197571231 | |
Розничная цена для студентов: 29 долларов.95 Элизабет А. Кларк 97801 237Мягкая обложка Февраль 2021 г. | |
Розничная цена для студентов: 19,95 $ Робин Уотерфилд 97801 765Мягкая обложка Март 2020 г. | |
Третье издание Розничная цена для студентов: 74 доллара.99 Ральф В. Мэтисен 97801945 Самый наводящий на размышления, современный и увлекательный обзор древнего мира | |
Четвертое издание Розничная цена для студентов: 54,99 $ Сара Б. Помрой, Стэнли М. Бурштейн, Уолтер Донлан… 97801 Наиболее сбалансированное освещение политической, военной, социальной, культурной и экономической истории Древней Греции | |
Розничная цена для студентов: 69 долларов.99 Ральф В. Мэтисен 97801 603 Сочетает увлекательное повествование и обширные первоисточники, чтобы сделать римскую историю более доступной, чем когда-либо прежде | |
Четвертое издание Розничная цена для студентов: 74,99 $ Сара Б. Померой, Стэнли М. Бурштейн, Уолтер Донлан. 97801918 Наиболее сбалансированное освещение политической, социальной, культурной и экономической истории Древней Греции | |
Розничная цена для студентов: 64,99 $ Ральф В. Мэтисен 97801918 Справочник, рассказывающий о цивилизациях и культурах древнего Средиземноморья | |
Розничная цена для студентов: 89 долларов.99 Фукидид 9780195002188 |
 Шульц
Шульц
 ..
..РЕКОНСТРУКЦИЯ НИКОЛА ВИСЕНТИНО ДРЕВНЕГРЕЧЕСКИХ РОДОВ на JSTOR
Информация о журнале La Revista de Musicología, выпущенный в 1978 году в качестве научного органа Sociedad Española de Musicología. Está dedicada especialmente al estudio del patrimonio music español y sus ramificaciones históricas y geográficas. Sus lineas de Investigacion abarcan todos los períodos y disciplinas, incluyendo artículos sobre música histórica, estética, teoría music, documentación, prácticaterpretativa, análisis y socialologia de la música. Incluye además уна sección Fija кон recensiones críticas де libros де reciente publicación. Revista de Musicología (Журнал музыковедения) был впервые опубликован в 1978 году и считается научным органом Испанского музыковедческого общества. Он в основном ориентирован на изучение испанского музыкального наследия и его различных географических и исторических ветвей.Его исследовательские направления охватывают все периоды и дисциплины, включая статьи об исторической музыке, эстетике, теории музыки, документации, игровой практике, анализе и социологии музыки. Он также содержит постоянный раздел с рецензиями на недавно изданные книги.
Sus lineas de Investigacion abarcan todos los períodos y disciplinas, incluyendo artículos sobre música histórica, estética, teoría music, documentación, prácticaterpretativa, análisis y socialologia de la música. Incluye además уна sección Fija кон recensiones críticas де libros де reciente publicación. Revista de Musicología (Журнал музыковедения) был впервые опубликован в 1978 году и считается научным органом Испанского музыковедческого общества. Он в основном ориентирован на изучение испанского музыкального наследия и его различных географических и исторических ветвей.Его исследовательские направления охватывают все периоды и дисциплины, включая статьи об исторической музыке, эстетике, теории музыки, документации, игровой практике, анализе и социологии музыки. Он также содержит постоянный раздел с рецензиями на недавно изданные книги.
La SOCIEDAD ESPAÑOLA de MUSICOLOGÍA (SEdeM) является научной, культурной и доцентской ассоциацией. Tiene por objeto el estudio y la enseñanza de la Musicología y de la música в целом, против особенного énfasis en el conocimiento, recuperación y difusión del patrimonio music español.Promueve toda clase de tareas musicológicas relacionadas con el mismo, tales como organizar congresos, simposios o jornadas de estudio, fomentar la Investigación musicológica y ayudar, en la manera posible, a la publicación de trabajos especializados. Испанское музыковедческое общество (SEdeM) является научной, культурной и образовательной ассоциацией. Его целью является изучение и преподавание музыковедения и музыки в целом, уделяя особое внимание познанию, восстановлению и распространению испанского музыкального наследия.Ассоциация спонсирует и организует множество музыковедческих мероприятий, таких как конференции или симпозиумы, и поощряет музыковедческие исследования, помогая публиковать как можно больше специализированных статей.
Tiene por objeto el estudio y la enseñanza de la Musicología y de la música в целом, против особенного énfasis en el conocimiento, recuperación y difusión del patrimonio music español.Promueve toda clase de tareas musicológicas relacionadas con el mismo, tales como organizar congresos, simposios o jornadas de estudio, fomentar la Investigación musicológica y ayudar, en la manera posible, a la publicación de trabajos especializados. Испанское музыковедческое общество (SEdeM) является научной, культурной и образовательной ассоциацией. Его целью является изучение и преподавание музыковедения и музыки в целом, уделяя особое внимание познанию, восстановлению и распространению испанского музыкального наследия.Ассоциация спонсирует и организует множество музыковедческих мероприятий, таких как конференции или симпозиумы, и поощряет музыковедческие исследования, помогая публиковать как можно больше специализированных статей.
Род Colchicum в трудах древнегреческих и византийских врачей
Обзор
дои: 10. 2174/1381612824666180115111546.Принадлежности Расширять
Принадлежности
- 1 Университетский институт истории медицины, медицинский факультет, Лионский университет им. Клода Бернара 1, Лион, Франция.
- 2 Отделение биомедицинской инженерии, первое отделение кардиологии, больница Гиппократион, медицинский факультет Афинского национального и Каподистрийского университета, Афины, Греция.
- 3 Кафедра фармакологии, Медицинский факультет, Патрский университет, Патры, Греция.
- 4 Лаборатория биологии опухолевых клеток, Медицинская школа, Критский университет, Ираклион, Греция.

- 5 Отделение кардиологии, Афинская больница общего профиля им. Г. Генниматаса, Афины, Греция.

Элемент в буфере обмена
Обзор
Грегори Цукалас и соавт. Курр Фарм Дез. 2018.
Показать детали Показать вариантыПоказать варианты
Формат АннотацияPubMedPMID
дои: 10.2174/1381612824666180115111546.Принадлежности
- 1 Университетский институт истории медицины, медицинский факультет, Лионский университет им. Клода Бернара 1, Лион, Франция.
- 2 Отделение биомедицинской инженерии, первое отделение кардиологии, больница Гиппократион, медицинский факультет Афинского национального и Каподистрийского университета, Афины, Греция.
- 3 Кафедра фармакологии, Медицинский факультет, Патрский университет, Патры, Греция.
- 4 Лаборатория биологии опухолевых клеток, Медицинская школа, Критский университет, Ираклион, Греция.
- 5 Отделение кардиологии, Афинская больница общего профиля им. Г. Генниматаса, Афины, Греция.
 Клода Бернара 1, Лион, Франция.
Клода Бернара 1, Лион, Франция.Элемент в буфере обмена
Полнотекстовые ссылки Параметры отображения цитированияПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Растения семейства безвременников были известны в архаический период в Греции своими вредными свойствами. Позже их использовали для лечения подагры. Лечение было введено древнегреческими врачами и передано византийским и арабским врачам и сохранилось до наших дней. Первое растение, скорее всего, было названо «Медея» в честь пресловутой колхидской ведьмы. Поскольку наиболее распространенный член семейства цветет осенью, растение было названо безвременником осенним. Также использовались различные номинации, такие как Эфемерон, Гермодактиль, Анима артикулорум и Суруген. Наша статья обсуждает их, и в то же время представляет наиболее известных авторитетов, которые использовали растения безвременника в фитотерапии и токсикологии.
Позже их использовали для лечения подагры. Лечение было введено древнегреческими врачами и передано византийским и арабским врачам и сохранилось до наших дней. Первое растение, скорее всего, было названо «Медея» в честь пресловутой колхидской ведьмы. Поскольку наиболее распространенный член семейства цветет осенью, растение было названо безвременником осенним. Также использовались различные номинации, такие как Эфемерон, Гермодактиль, Анима артикулорум и Суруген. Наша статья обсуждает их, и в то же время представляет наиболее известных авторитетов, которые использовали растения безвременника в фитотерапии и токсикологии.
Ключевые слова: безвременник; древнегреческая медицина; эфемерон; травяной медицины.; гермодактиль; токсикология..
Авторские права © Bentham Science Publishers; По любым вопросам пишите по адресу epub@benhamscience. org.
org.
Похожие статьи
- От яда к наркотику: обнаружены новые рецепты, содержащие колхицин как средство от подагры в Риме и Византии.
Урсин Ф., Стегер Ф. Урсин Ф. и соавт. Клин Эксперт Ревматол. 2019 июль-август;37(4):540-545. Epub 2018 19 ноября. Клин Эксперт Ревматол. 2019. PMID: 30557131
- Народная медицина и современность: терапия подагры.
Алушик Т, Алушик Ш. Алушик Т. и др. Кас Лек Ческ. 2017 Зима; 156(8):454-457. Кас Лек Ческ. 2017.PMID: 29368528 Чешский язык.
- Колхицин — краткая история древнего лекарства.
Нерлекар Н., Бил А., Харпер Р.В. Нерлекар Н. и соавт. Мед J Aust.
2014 11 декабря; 201 (11): 687-8. doi: 10.5694/mja14.00846.
Мед J Aust. 2014.
PMID: 25495322
Аннотация недоступна. - Выделение колхицина в 19 веке: новый взгляд на старый препарат.
Караману М., Цукалас Г., Пантос К., Андруцос Г. Караману М. и соавт. Курр Фарм Дез. 2018;24(6):654-658. дои: 10.2174/1381612824666180115105850. Курр Фарм Дез. 2018. PMID: 29336251 Обзор.
- Колхицин: обзор химической структуры и клинического использования.
Алкади Х., Хубейз М.Дж., Джбейли Р. Алкади Х. и др. Заразить мишени для наркотиков.2018;18(2):105-121. дои: 10.2174/1871526517666171017114901. Заразить мишени для наркотиков. 2018. PMID: 264 Обзор.
 2014 11 декабря; 201 (11): 687-8. doi: 10.5694/mja14.00846.
Мед J Aust. 2014.
PMID: 25495322
Аннотация недоступна.
2014 11 декабря; 201 (11): 687-8. doi: 10.5694/mja14.00846.
Мед J Aust. 2014.
PMID: 25495322
Аннотация недоступна.Цитируется
1 артикул- Перепрофилирование пути колхицина с учетом взаимодействий между лекарственными средствами. Обзор.
Врачатис Д.А., Папатанасиу К.А., Гиотаки С.Г., Илиодромитис К.Е., Папаиоанну Т.Г., Стефанини Г.Г., Клеман М., Сиасос Г., Реймерс Б., Лански А., Тардиф Дж.С., Дефтереос С.Г., Яннопулос Г.Врачатис Д.А. и соавт. Toxicol Rep. 9 июля 2021 г .; 8: 1389-1393. doi: 10.1016/j.toxrep.2021.07.009. Электронная коллекция 2021. Токсикол Респ. 2021. PMID: 34285885 Бесплатная статья ЧВК.
 Обзор.
Обзор.термины MeSH
- Колхицин / терапевтическое применение*
- Супрессоры подагры / история
- Супрессоры подагры / терапевтическое применение*
- Растения, Лекарственные / химия
LinkOut — больше ресурсов
Полнотекстовые источники
Прочие литературные источники
Медицинские
Исследовательские материалы
Укажите
КопироватьФормат: ААД АПА МДА НЛМ
Древние товары: Tria Genera Bonorum в этической теории
Страница из
НАПЕЧАТАНО ИЗ OXFORD SCHOLARSHIP ONLINE (oxford. Universitypressscholarship.com). (c) Copyright Oxford University Press, 2022. Все права защищены. Индивидуальный пользователь может распечатать PDF-файл одной главы монографии в OSO для личного использования. Дата: 04 февраля 2022 г.
Universitypressscholarship.com). (c) Copyright Oxford University Press, 2022. Все права защищены. Индивидуальный пользователь может распечатать PDF-файл одной главы монографии в OSO для личного использования. Дата: 04 февраля 2022 г.
Tria Genera Bonorum в этической теории
- Глава:
- (стр. 255) 11 Старинные товары
- Источник:
- Strategies of Argument
- Автор(ы):
Брэд Инвуд
- Издатель:
- Oxford University Press .1093/acprof:oso/97801998.003.0011
- Предусмотренная законом защита для захоронений старше 150 лет
- Регулярные консультации с индийской общиной через Индийский консультативный совет
- Экспертиза и исследование человеческих останков
- Повторное захоронение человеческих останков на специально отведенном государственном кладбище
- Публикация результатов исследования
- Общественное образование по охране мест захоронения и бережному отношению к местам захоронения и человеческим останкам
В этой главе в эллинистический период прослеживается использование древней классификации благ на три вида: блага тела, блага разума и «внешние» блага, т. е. блага, расположенные вне человека. Это деление берет свое начало у Платона и Аристотеля, его можно найти в отчетах о платоновской этике, в более поздней перипатетической этике, и оно оказало косвенное влияние на стоицизм. В нем делается вывод о том, что в эллинистический период поиск все более совершенных классификаций стал самоцелью.Тройственная классификация благ превратилась в доктрину, и чрезмерная озабоченность ею затуманивала обсуждение благ. Сенека, например, решительно и понятно выступает против одержимости стоиков категориями товаров.
В нем делается вывод о том, что в эллинистический период поиск все более совершенных классификаций стал самоцелью.Тройственная классификация благ превратилась в доктрину, и чрезмерная озабоченность ею затуманивала обсуждение благ. Сенека, например, решительно и понятно выступает против одержимости стоиков категориями товаров.
Ключевые слова: Античная этика, эллинистическая этика, Платон, Аристотель, стоицизм, tria genera bonorum, разделение благ, внешние блага
Oxford Scholarship Online требует подписки или покупки для доступа к полному тексту книг в рамках службы.Однако общедоступные пользователи могут свободно осуществлять поиск по сайту и просматривать рефераты и ключевые слова для каждой книги и главы.
Пожалуйста, подпишитесь или войдите, чтобы получить доступ к полнотекстовому содержимому.
Если вы считаете, что у вас должен быть доступ к этому названию, обратитесь к своему библиотекарю.
Для устранения неполадок см. Часто задаваемые вопросы , и если вы не можете найти ответ там, пожалуйста, связаться с нами .
Медицина и психиатрия в западной культуре: древнегреческие мифы и современные предрассудки | Annals of General Psychiatry
Давно замечено, что слоны и свиньи наслаждаются эффектами алкоголя, получаемого при употреблении в пищу опавших манго или ферментирующих яблок. Похоже, наши предки наблюдали и копировали поведение таких животных, а затем, будучи людьми и обладая способностью мыслить и творить, разработали методы, обеспечивающие непрерывный запас алкоголя.Можно предположить, что тогда, как и сейчас, ценностью такого процесса для выживания было стремление к временному избавлению от человеческого состояния.
Натуральные психотропные вещества, полученные из растений, включая табак, каннабис, опиум, кофе и другие, обеспечили необходимое спасение, составив так называемую «доалкогольную эру».
В то же время преследовалось как рекреационное, так и терапевтическое использование природных ресурсов. Глядя на свое ближайшее окружение, люди не различали вещества, предназначенные для лечения болезней или облегчения боли.Действительно, обе цели первоначально рассматривались как неделимые потребности и лишь позднее стали рассматриваться отдельно. Более того, соматические и психические заболевания изначально рассматривались как часть одного целого, поэтому и лечились с помощью одних и тех же веществ.
Психические и соматические симптомы рассматривались как отдельные явления только в более поздние времена, прежде чем, наконец, снова стали рассматриваться как различные проявления, воздействующие на «одно и то же тело» в ходе «одной и той же болезни».
Вместо того, чтобы искать лекарства в природных источниках, человеческая потребность искать лекарство от боли, болезней и смерти была решена путем обращения к сверхъестественному и магии. Неудивительно, что слово «лекарство» происходит от латинского глагола «mederi», что также напоминает латинское происхождение слова «лекарство». На самом деле, главная цель медицины состоит в том, чтобы излечить боль и болезнь независимо от их источника.
На самом деле, главная цель медицины состоит в том, чтобы излечить боль и болезнь независимо от их источника.
Древнегреческая медицина была сложной практикой, воспринимаемой как нечто среднее между мифом и реальностью, как выражение магической гадательной, иератической и эмпирической технической практики.Следовательно, античная медицина тесно связана с античной мифологией.
Примером такого совпадения является миф об Асклепии, которого древние греки считали, цитируя Пиндара (522–443 гг. до н. э.), «богом медицины» [8].
Согласно мифу, Асклепий, сын бога Аполлона и нимфы Корониды, родился от мертвого тела своей матери, неверной жены, казненной богиней Артемидой, сестры-близнеца Аполлона. Это считается первым кесаревым сечением от мертвой матери.Асклепий был воспитан Хироном, кентавром, опять же пересечением мифа и медицинских знаний, считавшимся мастером медицинской практики и лекарственных трав. Вскоре ученик превзошел своего учителя, став «богом медицины».
Гомер (около 8 века до н. э.) также сообщает, что Асклепий был первым, кто провел различие между медициной и хирургией: он дал силу выздоровления своему сыну Подалириусу и способность лечить раны Махаону; интересно, что никакого разделения между психическими и соматическими состояниями не проводилось [9].
э.) также сообщает, что Асклепий был первым, кто провел различие между медициной и хирургией: он дал силу выздоровления своему сыну Подалириусу и способность лечить раны Махаону; интересно, что никакого разделения между психическими и соматическими состояниями не проводилось [9].
Большинство древних мифов обычно связаны с событиями повседневной жизни. В Элладе было построено более 200 храмов Асклепия, представляющих собой первые известные больницы. Пациентам разрешалось отдыхать и спать рядом с аркадами, где снились пророческие сны, вероятно, вызванные невольным употреблением наркотиков. Они считали, что во время таких снов могут ощущать присутствие Асклепия, дающего им терапевтические советы и способность к выздоровлению. На самом деле, врачи-волшебники, одетые как боги, давали пациентам яды по ночам.
Храмы долгое время считались предпочтительным местом для магических и медицинских ритуалов, и животные часто присутствовали во время таких церемоний. Мифологический образ Корониды, матери бога медицины, также связан с терапевтическим ритуалом, совершаемым в священном храме Афины. Название Коронис происходит от древнегреческого слова «corònos», означающего «вороны», которое считается связанным с древним диагностическим и терапевтическим ритуалом, совершаемым для предсказания будущего.Во время этой практики можно было принять участие в перевоплощении героев в воронов или змей благодаря вмешательству богини Афины, также известной как «пользователь омелы».
Название Коронис происходит от древнегреческого слова «corònos», означающего «вороны», которое считается связанным с древним диагностическим и терапевтическим ритуалом, совершаемым для предсказания будущего.Во время этой практики можно было принять участие в перевоплощении героев в воронов или змей благодаря вмешательству богини Афины, также известной как «пользователь омелы».
Сбор омелы, растущей на ветвях дуба, сохранил символическое значение кастрации дерева-хозяина, потому что сок из ягод омелы, рассматриваемый как «дубовая сперма» (омела называлась «Viscum Album» на латыни, возможно, из-за ее спермоподобных свойств ) считалось заряженным регенеративными способностями и терапевтическими свойствами [10].Слово «омела» также связано с латинским именем бога медицины Асклепия, означающего «то, что свисает со съедобного дуба» или «эскулент», то есть «хорошее для еды». омела также означает.
Любопытно отметить, что в разных цивилизациях и в разные эпохи одни и те же вещества использовались с одинаковыми целями, тогда как сегодня большинство этих значений почти забыто. Например, кельты тоже считали омелу священной, используя ее во многих религиозных ритуалах.Это растение также широко применялось как лечебное средство в Средние века и в эпоху Возрождения, а ко второй половине 19-го – первому десятилетию 20-го века его стали назначать из-за его антигипертензивных свойств. Сегодня он имеет просто символическое значение на Рождество.
Например, кельты тоже считали омелу священной, используя ее во многих религиозных ритуалах.Это растение также широко применялось как лечебное средство в Средние века и в эпоху Возрождения, а ко второй половине 19-го – первому десятилетию 20-го века его стали назначать из-за его антигипертензивных свойств. Сегодня он имеет просто символическое значение на Рождество.
Омела — лишь одно из множества различных растений и деревьев, использовавшихся в древней медицинской практике; кора ив, окружающих храм Афины, содержала салицилат, жаропонижающее и противовоспалительное средство, до сих пор широко назначаемое современными врачами (сегодня известно, что оно способно взаимодействовать с реактивным провоспалительным С-белком).
Независимо от мифов, используемых для объяснения болезней и их лечения, совпадения между легендами, религией и медициной были широко распространены среди древних греков, пока Гиппократ (460–377 г. до н. э.) впервые не выделил такие сущности.
Гиппократ представил более современную модель депрессии. Он применил концепции своей основной «теории четырех дош и темпераментов» к меланхолии («мелайна» означает «черный», а «холе» — «желчь»), описанной как следствие дисбаланса четырех основных жидкостей, влияющих на оба физических состояния. и психические проявления.Этими жидкостями («гуморами») были кровь, мокрота, черная желчь и желтая желчь.
Что касается психических расстройств, то флюидная теория в равной степени применима и к соматическим проявлениям. Согласно теории Гиппократа, дисбаланс в пропорциях жидкости считался ответственным за своеобразный темперамент, который, возможно, приводил к тяжелой депрессии, в то время как у здорового человека, человека с «хорошим чувством юмора», четыре жидкости находятся в соответствующих пропорциях. Таким образом, депрессия и другие психические состояния считались органическими расстройствами.
Современным врачам известно, что при течении рака поджелудочной железы у большинства больных возникает тяжелая «меланхолическая» депрессия, почти неотличимая от первичной депрессии; древние врачи уже отмечали это клиническое явление, и, вероятно, поэтому поджелудочную железу считали внутренним местопребыванием черной желчи.
Используя вышеупомянутую теорию, Гиппократ был также первым, кто описал возможные сезонные течения расстройств «настроения» или «юмора» [11]. Большинство наблюдений Гиппократа представляли собой новаторские достижения в области медицины.Тем не менее, они обычно игнорировались или «неправильно понимались» в последующие века, когда миф (или религия) влиял на медицинский и научный подход.
Защита древних человеческих останков в Айове — общая информация
Программа биоархеологии
Управление государственного археолога
Университет Айовы
Индийский консультативный совет OSA
Сюзанна Ванати Баффало (Мескваки)
Лэнс Фостер (Айова)
Джеки Рэнд (чокто)
Джо Коултер (Потаватоми)
Джули Мартино (Лакота)
Марк Рэйвенхейр (Оглала Сиу)
Джон Ф. Доершук, государственный археолог
Доершук, государственный археолог
Лара Нольднер, директор по биоархеологии
Программа Айовы по защите древних человеческих останков
Защита человеческих останков в Айове
Управление государственного археолога (OSA) является подразделением Университета Айовы.В его обязанности входит исследование, интерпретация и сохранение древних захоронений, а также, при необходимости, восстановление и перезахоронение останков древних людей. Закон штата возложил эти обязанности на OSA в 1976 году после того, как индейцы в Айове подняли вопросы надлежащего распоряжения индейскими захоронениями, осквернения индейских могильников и равной защиты в соответствии с законом. Принятая совместными усилиями представителей Индии, археолога штата, законодателей штата и губернатора, глава 263B Кодекса Айовы защищает останки древних людей в Айове.OSA тесно сотрудничает с индийской общиной через Индийский консультативный совет для реализации закона.
Закон штата возложил эти обязанности на OSA в 1976 году после того, как индейцы в Айове подняли вопросы надлежащего распоряжения индейскими захоронениями, осквернения индейских могильников и равной защиты в соответствии с законом. Принятая совместными усилиями представителей Индии, археолога штата, законодателей штата и губернатора, глава 263B Кодекса Айовы защищает останки древних людей в Айове.OSA тесно сотрудничает с индийской общиной через Индийский консультативный совет для реализации закона.
Древние человеческие останки старше 150 лет. Правовая защита захоронений в Айове распространяется на доисторические курганы и безымянные кладбища. Сайты сохраняются и защищаются, когда это возможно. Однако, если человеческие останки были или должны быть удалены, OSA проводит эксгумацию и исследование останков перед повторным захоронением.
Любой человек, обнаруживший возможное место древнего захоронения или скелетный материал, который, как он или она подозревает, может быть человеком, должен связаться с OSA.
Если будет определено, что кости принадлежат человеку, они будут исследованы физическим антропологом в попытке определить культурную принадлежность. Отчет будет написан и представлен в Департамент общественного здравоохранения штата Айова. Если будет установлено, что останки принадлежат коренным американцам и, следовательно, подпадают под действие Закона о защите и репатриации могил коренных американцев (NAGPRA), будут проведены надлежащие процессы уведомления и консультаций.
Любой, кто планирует строительные работы и беспокоится о захоронениях, должен обратиться в OSA за помощью, чтобы обеспечить соблюдение закона Айовы и сохранить наше невозобновляемое культурное наследие.Любой, кто знает о нарушении древних человеческих останков, должен связаться с OSA. Умышленное нарушение захоронения считается преступным хулиганством третьей степени (глава 716.5 Кодекса Айовы).
OSA готово более подробно объяснить процедуры, применяемые при обнаружении человеческих останков, что мы можем узнать из костей, а также важность уважительного обращения с человеческими останками и захоронениями. Пожалуйста, свяжитесь с OSA для получения дополнительной информации.
Пожалуйста, свяжитесь с OSA для получения дополнительной информации.
Биоархеологическая программа OSA
Биоархеологическая программа OSA участвует в многочисленных полевых проектах по всей Айове.Музеи, другие хранилища и частные лица также представляют свои коллекции в OSA для изучения и перезахоронения. С 1976 года в рамках программы реализовано более 3000 проектов во всех 99 округах штата Айова.
Полевые проекты приводят к открытию множества новых участков и получению новой информации о ранее зарегистрированных участках. Полевые работы включают проверку сообщений о возможных захоронениях или курганах, а также осмотр курганов или групп курганов и других захоронений, которым угрожает эрозия, строительные или карьерные работы или вандализм.Многие сайты требуют периодического мониторинга на постоянной основе. Сохранение сайтов всегда является предпочтительным лечением. В случаях, когда важные объекты находятся под угрозой, государственный археолог имеет право (Кодекс штата Айова, 263B. 9) отказать в разрешении на изъятие.
9) отказать в разрешении на изъятие.
Результаты анализов человеческих останков публикуются в целях распространения информации, полученной в соответствии с уставным обязательством OSA по публикации образовательных и научных отчетов. Отчеты Программы биоархеологии, которые подаются в Департамент общественного здравоохранения штата Айова, объединяют восстановленную культурную и физическую информацию и служат для установления значимости расследуемых объектов.С начала программы были перезахоронены останки более 2000 человек. Для этой цели в 1977 году было открыто и выделено кладбище на государственной земле. Второе кладбище было создано в 1979 году, третье — в 1987 году и четвертое — в 1991 году. Останки повторно захоронены на кладбище, ближайшем к первоначальному месту захоронения. и церемония проводится во время или вскоре после каждого перезахоронения в соответствии с пожеланиями Индийского консультативного совета. Репатриация культурно связанных останков осуществляется в соответствии с NAGPRA.
Биоархеологическая программа OSA посвящена защите захоронений и человеческих останков, перезахоронению древних человеческих останков и более глубокому пониманию прошлого Айовы. Регулярные консультации с Индийским консультативным советом OSA являются важным элементом программы. Все жители Айовы обслуживаются в рамках программы защиты наследия Айовы.
«В религии Мескаки наш создатель поместил нас на эту землю (не в Европу или Африку, а сюда).Земля — главное хранилище для всех людей… Мескаки приспособились к современным обычаям, но наши захоронения не изменились. Все захоронения остаются прежними, неприкосновенными».
Дон Ванати, 1980 г. Семинар по планированию древних могильников.
«Индийская точка зрения на эксгумацию может быть изложена простыми словами. В основном редко бывает веская причина для извлечения мертвых из земли, где они так долго покоились, а когда возникает необходимость, эти человеческие останки должны быть перезахоронены как как можно скорее и надлежащим и достойным образом. … Наши предки были людьми, и мы хотим, чтобы их оставили в покое… Мы, индейцы, уважаем наших предков. Они присутствуют на наших церемониях, и мы призываем их на помощь, чтобы прожить нашу жизнь, помогая друг другу … [Они] когда-то были людьми, которые когда-то ходили по земле, любили, молились, заботились о своих семьях, умерли и были похоронены. … Мы считаем, что мертвые должны оставаться похороненными. Земли достаточно для всего, что нужно сделать, так что несколько акров, оставленных для наших мертвецов, не заставят никого голодать.»
… Наши предки были людьми, и мы хотим, чтобы их оставили в покое… Мы, индейцы, уважаем наших предков. Они присутствуют на наших церемониях, и мы призываем их на помощь, чтобы прожить нашу жизнь, помогая друг другу … [Они] когда-то были людьми, которые когда-то ходили по земле, любили, молились, заботились о своих семьях, умерли и были похоронены. … Мы считаем, что мертвые должны оставаться похороненными. Земли достаточно для всего, что нужно сделать, так что несколько акров, оставленных для наших мертвецов, не заставят никого голодать.»
Мария Пирсон, статья под названием «Индийская точка зрения: эксгумация и перезахоронение».
«За столетия в Айове погибло больше индейцев, чем кто-либо или какой-либо народ сегодня может когда-либо вспомнить имена. Исчезли целые племена, не оставив своих имен нашему собственному поколению… Я говорю, что возможно, увидеть некоторые из эти старожилы как живые, дышащие, мыслящие, чувствующие личности с верованиями, страхами, надеждами и молитвами. В этом смысле эти люди на самом деле не мертвы. Если мы можем видеть их как реальных людей, они на самом деле не мертвы.»
В этом смысле эти люди на самом деле не мертвы. Если мы можем видеть их как реальных людей, они на самом деле не мертвы.»
Роберт Л. Холл, антрополог, Симпозиум 1983 года по изучению скелетных останков древних людей в Айове.
«У индийцев и ученых есть много веских причин для защиты и сохранения древних захоронений. Нам нужно сформировать партнерство и работать вместе для достижения общих целей. Мы союзники, и мы должны работать бок о бок против сил эрозии, развития, грабежей и других факторов, уничтожающих древние могильники.
Дуэйн Андерсон, бывший государственный археолог, 1980 г. Семинар по планированию древних могильников.
.